



ņä£ļĪĀ
ņŚ╝ ņŖżĒŖĖļĀłņŖżļŖö ņé╝Ēł¼ņĢĢņ£╝ļĪ£ ņØĖĒĢ£ ņØ┤ņś© ļČłĻĘĀĒśĢ ļ░Å ļ┐īļ”¼ņØś ņłśļČäĒØĪ ņłś ņĀĆĒĢś ļ┐É ņĢäļŗłļØ╝ ņØ┤ņ░©ņĀü ņé░ĒÖöņŖżĒŖĖļĀłņŖżļź╝ ļÅÖļ░śĒĢśļŖö ļ│ĄĒĢ®ņĀü ņŖżĒŖĖļĀł ņŖż ņÜöņØĖ ņżæņØś ĒĢśļéśņØĖļŹ░, ņŗØļ¼╝ņ▓┤ļŖö ņŚ╝ ņŖżĒŖĖļĀłņŖż ņŗ£ ņłśļ░śļÉśļŖö ĒÖ£ņä▒ņé░ ņåīņóģņØś ņ”ØĻ░Ćļź╝ ļ¦ēĻĖ░ ņ£äĒĢ┤ ĒĢŁņé░ĒÖö ļ¼╝ņ¦ł ļ░Å ĒĢŁņé░ĒÖö ĒÜ©ņåīļōżņØä ņāØņé░ĒĢ£ ļŗż. Kim et al.(2010)ņØĆ ņĀĆ ļåŹļÅäņØś ņŚ╝ņ▓śļ”¼Ļ░Ć ļ░░ņČöņØś ņāØņ£Ī ļ░Å ĒÆłņ¦łņØä ņ┤ēņ¦äņŗ£ņ╝░ļŗżĻ│Ā ļ│┤Ļ│ĀĒĢ£ ļ░ö ņ׳ļŗż. ĻĘĖļ¤¼ļéś ņŚ╝ ņŖżĒŖĖļĀłņŖż ņŗ£ ņ”ēĻ░üņĀüņØ┤Ļ│Ā ņØ╝ņŗ£ņĀüņ£╝ļĪ£ ņ”ØĻ░ĆĒĢśļŖö ĒĢŁņé░ĒÖö ĒÜ©ņåīņÖĆļŖö ļŗ¼ļ”¼, ĒĢŁņé░ĒÖö ļ¼╝ņ¦łļōżņØĆ 2ņ░© ļīĆņé¼ņé░ļ¼╝ļĪ£ ņāØņä▒ļÉ£ Ēøä ņŗØļ¼╝ņ▓┤ ļé┤ņŚÉ ņČĢņĀüļÉ£ļŗż(Ross and Sombrero, 1991; Delfine et al., 2005). ĒĢŁņé░ĒÖöņä▒ņØä ļéśĒāĆļé┤ļŖö ņŗØļ¼╝ņØś 2ņ░©ļīĆņé¼ ņé░ļ¼╝ņØĆ ņŚ╝ ņŖżĒŖĖļĀłņŖż ņÖĖņŚÉ Ļ│Āņś©, Ļ▒┤ņĪ░, Ļ░ĢĻ┤æ ļō▒Ļ│╝ Ļ░ÖņØĆ ĒÖśĻ▓Į ņŖżĒŖĖļĀłņŖż ņÜöņØĖņØä ĒżĒĢ©ĒĢ£ ņÖĖļČĆņ×ÉĻĘ╣ņŚÉ ļīĆĒĢ┤ ņ×ÉņŗĀņØä ļ│┤ĒśĖĒĢśĻĖ░ ņ£äĒĢ┤ ņāØņä▒ļÉśļŖö ļ¼╝ ņ¦łļĪ£ ņĢīļĀżņĀĖ ņ׳ļŗż(Munne-Bosch et al., 1999). ļīĆĒæ£ņĀüņØĖ ĒĢŁņé░ĒÖöļ¼╝ ņ¦łļĪ£ļŖö ascorbic acid(ļ╣äĒāĆļ»╝ C), ╬▒-tocopherol(ļ╣äĒāĆļ»╝ E), ╬▓- carotene, glutathione(GSH), ubiquinone, ņÜöņé░ ļō▒ņØś ņĀĆļČäņ×É ĒĢŁņé░ ĒÖöļ¼╝ņ¦łĻ│╝ ņ┤Ø ĒÅ┤ļ”¼ĒÄśļåĆļźś ļ░Å ņ┤Ø ĒöīļØ╝ļ│┤ļģĖņØ┤ļō£ ļō▒ņØä ļōż ņłś ņ׳ļŗż(Su et al., 1988; Alsher and Hess, 1993; Last et al., 1997). ņŗØļ¼╝ ņ£Āļל ņ▓£ ņŚ░ļ¼╝ ĒĢŁņé░ĒÖöņĀ£ļĪ£ ļ¦ÄņØĆ Ļ┤Ćņŗ¼ņØä ļ░øĻ│Ā ņ׳ļŖö ĒÄśļåĆņä▒ ĒÖöĒĢ®ļ¼╝ņØĆ ņøÉļל photodamageļĪ£ļČĆĒä░ ņ×ÉņŗĀņØä ļ│┤ĒśĖĒĢśĻ│Ā(Close and Mcarther, 2002), ņ┤łņŗØļÅÖļ¼╝Ļ│╝ ĒĢ┤ņČ®ļźśņŚÉ ļīĆĒĢ£ ļ░®ņ¢┤ĻĖ░ņ×æņ£╝ļĪ£ ņāØņä▒, ņČĢņĀüļÉśļŖö ņä▒ ļČäņ£╝ļĪ£ ņĢīļĀżņĀĖ ņ׳ļŗż(Stevenson et al., 1993).
ĒŚłļĖīņŗØļ¼╝ņØś ņĀĢņ£Ā ņä▒ļČä ņāØĒĢ®ņä▒ ņŚŁņŗ£ ņ£ĀņĀäņĀü ņÜöņØĖ(Hay and Waterman, 1993) ņÖĖņŚÉ ĒÖśĻ▓ĮņĀü ņÜöņØĖņØś ņśüĒ¢źņØä ļ░øļŖöļŗż. ņĀĢņ£Ā ņłśņ£© ļ░Å ĒÆłņ¦łņØĆ ņ×¼ļ░░ņ¦ĆņŚŁ ļ░Å ņ¦łņåī ĒśĢĒā£(Shin and Park, 1999; Choi et al., 2010), ņ×¼ļ░░ļ░®ļ▓Ģ(Naiem et al., 1987; Mauro et al., 1994; Park et al., 1999), ņ▒äņĘ©ņŗ£ĻĖ░(Kim, 2005) ļō▒ņŚÉ ļö░ļØ╝ ņ░©ņØ┤Ļ░Ć ļé£ļŗż. ņØ┤ ļŖö ņĀüņĀłĒĢ£ ņŖżĒŖĖļĀłņŖż ņ▓śļ”¼Ļ░Ć ņśżĒ׳ļĀż ņŗØļ¼╝ņØś ļ░®ņ¢┤ ĻĖ░ņ×æņØä ņ×ÉĻĘ╣ĒĢśņŚ¼ ņĀĢ ņ£ĀņØś ņŻ╝ņÜö ĻĄ¼ņä▒ņä▒ļČäņØĖ terpenoidļéś ĒĢŁņé░ĒÖöņä▒ņØä ļéśĒāĆļé┤ļŖö phenolic compound ļō▒ņØś ĒĢ©ļ¤ē ņ”Øņ¦äņŚÉ ņśüĒ¢źņØä ļü╝ņ╣śĻ│Ā, ņĢäņÜĖļ¤¼ Ļ▓ĮņĀ£ņĀü Ļ│ĀļČĆ Ļ░Ć Ļ░Ćņ╣śņé░ļ¼╝ņØś ņ░ĮņČ£ Ļ░ĆļŖźņä▒ņØä ņŗ£ņé¼ĒĢ£ļŗż.
Ļ┐ĆĒÆĆĻ│╝(Lamiaceae) ĒŚłļĖīņŗØļ¼╝ņØś phenolic diterpenes(Kivilomopolo and Hyotylainen, 2007), flavonoid(Madsen and Bertelsen, 1995), phenolic acid(Cao and Cao, 1999) ļō▒ņØĆ ĒĢŁņé░ĒÖöņä▒ļ┐Éļ¦ī ņĢä ļŗłļØ╝ ĒĢŁĻĘĀņØä ļéśĒāĆļé┤ļŖö 2ņ░© ļīĆņé¼ņé░ļ¼╝ļĪ£ņä£, ņ▓£ņŚ░ ĒĢŁņé░ĒÖöņĀ£ļĪ£ņä£ņØś Ļ░£ ļ░£ Ļ░ĆļŖźņä▒ņØ┤ ļåÆņØĆ ņä▒ļČäļōżņØ┤ļŗż(Choudhury et al., 2006). Ļ┐ĆĒÆĆĻ│╝ ņŗØ ļ¼╝ņØś ļīĆĒæ£ņĀü ĒŚłļĖīņŗØļ¼╝ļĪ£ Ļ╝ĮĒ׳ļŖö ļ»╝ĒŖĖļŖö ņĢĮ 40ņŚ¼ņóģņØ┤ ņĢīļĀżņĀĖ ņ׳ļŖö ļŹ░, Ļ▓ĮņĀ£ņĀü Ļ░Ćņ╣śĻ░Ć Ēü░ ņŗØļ¼╝ņØ┤ņ¦Ćļ¦ī ņ×¼ļ░░ņŚÉ Ļ┤ĆĒĢ£ ņŚ░ĻĄ¼ļŖö ļ¦ÄņØ┤ ļČĆņĪ▒ĒĢś ļŗż(Diaz-maroto et al., 2003; Park, 2003). ĒŖ╣Ē׳ ņŚ╝ļźśņ¦æņĀü ļ░Å ņé░ņŚģ ļČĆņé░ļ¼╝ņØś ņ£Āņ×ģņ£╝ļĪ£ ņØĖĒĢ£ Ļ│ĀņŚ╝ļźś ĒåĀņ¢æ ņĪ░Ļ▒┤ņŚÉņä£ņØś ņ×¼ļ░░ļ▓Ģ ņŚ░ĻĄ¼ņÖĆ ņØ┤ ņÖĆ Ļ┤ĆļĀ©ĒĢ£ ĻĖ░ļŖźņä▒ĒāÉņāē ņŚ░ĻĄ¼ļÅä ĒĢäņÜöĒĢ£ ņŗżņĀĢņØ┤ļŗż(Amer, 1995). ļö░ļØ╝ ņä£ ļ│Ė ņŗżĒŚśņØĆ Ļ▓ĮņĀ£ņĀü Ļ░Ćņ╣śĻ░Ć ļåÆņØä ļ┐É ņĢäļŗłļØ╝ ĒĢŁĻĘĀ┬ĘĒĢŁņé░ĒÖöņä▒ņØ┤ ļåÆļŗż Ļ│Ā ļ│┤Ļ│ĀļÉ£ ļ»╝ĒŖĖļź╝ ņØ┤ņÜ®ĒĢśņŚ¼ ņŚ╝ ņŖżĒŖĖļĀłņŖż ņ▓śļ”¼ ņŗ£ ĒÆłņóģĻ░äņØś ņāØņ£Ī ļ░Å ņāØļ”¼ņĀü ļīĆņé¼ņ×æņÜ®Ļ│╝ ņŚ╝ņŚÉ ļīĆĒĢ£ Ļ░Éņłśņä▒ņØ┤ļéś ņĀĆĒĢŁņä▒ņØä ļ╣äĻĄÉĒĢśĻ│Āņ×É ņŗż ņŗ£ĒĢśņśĆļŗż. ņØ┤ļŖö ņĢ×ņ£╝ļĪ£ Ļ│ĀĻĖ░ļŖźņä▒ ņ×æļ¼╝ņØś ņĢłņĀĢņĀü ņ×¼ļ░░ņŗ£ņŖżĒģ£ Ļ░£ļ░£ ļ░Å ņé░ņŚģņĀü ņØæņÜ® Ļ░ĆļŖźņä▒ ĒÖĢļ│┤ņŚÉ ĒÖ£ņÜ®ņØ┤ Ļ░ĆļŖźĒĢĀ Ļ▓āņØ┤ļŗż.
ņŚ░ĻĄ¼ļ░®ļ▓Ģ
Ļ│Ąņŗ£ņ×¼ļŻīļŖö ĒÄśĒīīļ»╝ĒŖĖ(peppermint, Mentha ├Ś piperita), ņśżļŹ░ ņĮöļĪ▒ļ»╝ĒŖĖ(Eau de cologne mint, Mentha ├Ś piperita f. citrata), ņ┤ł ņĮöļ»╝ĒŖĖ(chocomint, Mentha ├Ś piperita f. citrata 'Chocolate'), ņŖż Ēö╝ņĢäļ»╝ĒŖĖ(spearmint, Mentha spicata), ņĢĀĒöīļ»╝ĒŖĖ(applemint, Mentha suaveolens), ĒīīņØĖņĢĀĒöīļ»╝ĒŖĖ(pineapplemint, Mentha suaveolens 'Variegata') ļō▒ ļ»╝ĒŖĖļźś 6ĒÆłņóģņØä ņé¼ņÜ®ĒĢśņśĆļŗż. ĒŚłļĖīļåŹņן ņØĖ ĒŚłļĖīļŗżņä»ļ®öņŚÉņä£ ņéĮļ¬®ļ¼śļź╝ ĻĄ¼ņ×ģĒĢśņŚ¼ ņé¼ņÜ®ĒĢśņśĆņ£╝ļ®░, ņŗżĒŚśņØĆ ņä£ņÜĖ ņŚ¼ļīĆ ļČĆņåŹļåŹņןņŚÉņä£ ņŗżņŗ£ĒĢśņśĆļŗż. ĒöīļØ╝ņŖżĒŗ▒ ĒĢśņÜ░ņŖżņØś ņś©ļÅäļŖö 25┬▒ 5┬░C, ņāüļīĆņŖĄļÅäļŖö 60-70%, ĒÅēĻĘĀņØ╝ņן 15ņŗ£Ļ░ä, ļ¦æņØĆ ļéĀ ņśżņĀä 10-12 ņŗ£Ļ▓ĮņŚÉ ņĖĪņĀĢĒĢ£ Ļ┤æĒĢ®ņä▒ņ£ĀĒÜ©Ļ┤æņ¢æņ×ÉņåŹņØĆ 330-370╬╝molŌłÖm-2ŌłÖs-1ņśĆļŗż. ņŗØļ¼╝ņ▓┤ļŖö ĒÅēĻĘĀ ņ┤łņן 10┬▒1cm, ņ×Ä 5ņןņØś ĻĘĀņØ╝ĒĢ£ Ļ░£ņ▓┤ļź╝ ņäĀļ░£ĒĢśņŚ¼ ĒÄä ļØ╝ņØ┤ĒŖĖļź╝ ņ▒äņÜ┤ ņ¦üĻ▓Į 10cm, ļåÆņØ┤ 10cmņØś ĒżĒŖĖņŚÉ ņØ┤ņŗØĒĢśņŚ¼ 2ņŻ╝Ļ░äņØś ļ┐īļ”¼ņØś ĒÖ£ņ░® ļ░Å ņĢłņĀĢ ĻĖ░Ļ░äņØä Ļ▒░ņ╣£ Ēøä ņŚ╝ ņ▓śļ”¼ļź╝ ņŗżņŗ£ĒĢśņśĆļŗż.
ņŚ╝ ņ▓śļ”¼ļŖö ņśłļ╣äņŗżĒŚśņØä Ļ▒░ņ│É ņłśļÅŚļ¼╝ ņ▓śļ”¼ĻĄ¼ņØĖ ļīĆņĪ░ĻĄ¼ļź╝ ĻĖ░ņżĆņ£╝ ļĪ£, ņłśļÅŚļ¼╝ņŚÉ 50, 100 ļ░Å 200mMņØś NaCl(Duchefa, 99%)ņØä ņ▓śļ”¼ ĒĢ£ 4ņ▓śļ”¼ĻĄ¼ļĪ£ ĻĄ¼ļČäĒĢśņśĆĻ│Ā, ņŻ╝ 2ĒÜī ĒżĒŖĖļŗ╣ 50mLņØś ņŚ╝ņ▓śļ”¼ ņÜ®ņĢĪņØä Ļ┤ĆņłśĒĢśņśĆļŗż. ņĢäņÜĖļ¤¼ hyponex ņĢĪļ╣ä(ņ¦łņåī 5%, ņłśņÜ®ņä▒ņØĖņé░ 10%, ņłś ņÜ®ņä▒Ļ░Ćļ”¼ 5%, ņØ╝ņłśĒÖöĒĢÖ ņĀ£ņĪ░) 1000ļ░░ ĒؼņäØņĢĪ 50mLļź╝ ņłśĒÖĢ ņŗ£Ļ╣ī ņ¦Ć ņŻ╝ 2ĒÜī Ļ┤ĆņłśĒĢśņśĆņ£╝ļ®░, ņ▓śļ”¼ĻĄ¼ļŗ╣ 5ĒżĒŖĖļź╝ ĒĢ£ ļ░śļ│Ąņ£╝ļĪ£ ĒĢśņŚ¼ 3ļ░ś ļ│Ą ņ▓śļ”¼ĒĢśņśĆļŗż.
ņŚ╝ ņ▓śļ”¼ 3ņŻ╝ Ēøä ņ┤łņן, ņŚĮņן, ņŚĮĒÅŁ, ņāØņ▓┤ņżæ ļ░Å Ļ▒┤ļ¼╝ļźĀņØä ņĪ░ņé¼ĒĢśņśĆ ļŗż. ņ┤łņןņØĆ ņ¦ĆņāüļČĆņÖĆ ņ¦ĆĒĢśļČĆņØś Ļ▓ĮĻ│äļĪ£ļČĆĒä░ Ļ░Ćņן ĻĖ┤ ņżäĻĖ░ņØś ņĀĢļŗ©Ļ╣ī ņ¦ĆņØś ĻĖĖņØ┤ļź╝ ņĖĪņĀĢĒĢśņśĆĻ│Ā, ņŚĮņןĻ│╝ ņśåĒÅŁņØĆ ņāüļŗ© 3ļ¦łļööņØś ņ×ÄņØä ļīĆņāüņ£╝ ļĪ£ ņ▓śļ”¼ĻĄ¼ ļé┤ņŚÉņä£ ĒÅēĻĘĀņĀüņØĖ Ēü¼ĻĖ░ņØś ņ×ÄņØś ĻĖĖņØ┤ņÖĆ ĒÅŁņØä ņĖĪņĀĢĒĢśņśĆļŗż. ņłśĒÖĢĒĢ£ ņŗØļ¼╝ņØĆ ņ¦ĆņāüļČĆņÖĆ ņ¦ĆĒĢśļČĆļĪ£ ĻĄ¼ļČäĒĢśņŚ¼ Ļ░üĻ░ü ņĀĢļ¤ēĒĢśĻ│Ā, 70┬░C Ļ▒┤ņĪ░ĻĖ░ņŚÉņä£ 1ņŻ╝ņØ╝Ļ░ä Ļ▒┤ņĪ░ņŗ£Ēé© Ēøä ļ¼┤Ļ▓īļź╝ ņĖĪņĀĢĒĢśņŚ¼ ņāØņ▓┤ņżæņŚÉ ļīĆĒĢ£ ļ░▒ļČäņ£©ļĪ£ ĒÖśņé░ĒĢśņśĆļŗż. ņŚĮļĪØņåī ņĖĪņĀĢņØĆ ņŚĮļĪØņåī ņĖĪņĀĢĻĖ░(SPAD 502, Minolta, Japan)ļź╝ ņé¼ņÜ®ĒĢśņŚ¼ ņĖĪņĀĢĒĢśņśĆļŗż. ņāØļ”¼ņĀü ļ░śņØæ ļ¼╝ņ¦łņØä ļČäņäØ ĒĢśĻĖ░ ņ£äĒĢ£ ņŗ£ļŻīļŖö ņŚ╝ ņ▓śļ”¼ 3ņŻ╝ĒøäņŚÉ ņĄ£ņóģ ņłśĒÖĢĒĢ£ ņŗØļ¼╝ņ▓┤ļź╝ ņ▒äņĘ©ĒĢśņŚ¼ ņé¼ņÜ®ĒĢśņśĆļŗż. DPPH(1,1-diphenyl-2- picrylhydrazyl) ļØ╝ļööņ╗¼ ņåī Ļ▒░ĒÖ£ņä▒ņØĆ Blios(1958)ņØś ļ░®ļ▓ĢņØä ņØæņÜ®ĒĢśņŚ¼ ņé¼ņÜ®ĒĢśņśĆĻ│Ā, ņ┤Ø ĒÅ┤ļ”¼ĒÄś ļåĆĒĢ©ļ¤ēņØĆ Folin-Denisļ▓Ģ(Folin and Denis, 1915)ņØä ņØæņÜ®(Velioglu, 1998)ĒĢśņŚ¼ ņĖĪņĀĢĒĢśņśĆņ£╝ļ®░, ņ┤Ø ĒöīļØ╝ļ│┤ļģĖņØ┤ļō£ ĒĢ©ļ¤ēņØĆ diethylene glycol ļ╣äņāēļ▓Ģ(NFRI, 1990)ņØä ņé¼ņÜ®ĒĢśņśĆļŗż. ĒåĄĻ│äņ▓śļ”¼ļŖö SPSSļź╝ ņØ┤ņÜ®ĒĢśņśĆņ£╝ļ®░, Ļ░ü ĻĄ░ņŚÉ ļö░ļźĖ ņ£ĀņØśņ░© Ļ▓ĆņĀĢņØĆ ļČäņé░ļČäņäØņØä ĒĢ£ Ēøä 5% ņłśņżĆņŚÉņä£ Duncan's multiple range testļź╝ ņŗżņŗ£ĒĢśņśĆļŗż.
Ļ▓░Ļ│╝ ļ░Å Ļ│Āņ░░
ņŚ╝ ņ▓śļ”¼ņŚÉ ļö░ļźĖ ļ»╝ĒŖĖ 6ĒÆłņóģņØś ņāØņ£Ī ļ░Å ņāØļ”¼ņĀü ļ░śņØæņØä ļ│┤ļ®┤ ņ┤łņן, ņŚĮņן, ņŚĮĒÅŁ ļō▒ ņÖĖĻ┤ĆņāüņØś ņāØņ£ĪņØĆ ņŚ╝ ļåŹļÅäĻ░Ć ņ”ØĻ░ĆĒĢĀņłśļĪØ Ļ░ÉņåīĒĢśņśĆĻ│Ā, ļåŹļÅäņŚÉ ļö░ļźĖ Ļ░Éņåīņ£©ņØĆ ĒÆłņóģ ļ░Å ļČĆņ£äņŚÉ ļö░ļØ╝ ņ░©ņØ┤Ļ░Ć ļéśĒāĆļé¼ļŗż(Table 1). Shim et al.(1998) ņŚŁņŗ£ ņŗØļ¼╝ņØś ņŚ╝ ņ╣śņé¼ļåŹļÅä, ņāØņן Ļ░Éņåīņ£©ņØĆ Ļ░Ö ņØĆ ņóģ Ļ░äņŚÉļÅä Ēü¼Ļ▓ī ņ░©ņØ┤Ļ░Ć ļéśņä£ ļé┤ņŚ╝ņä▒ ĒÆłņóģĻ│╝ Ļ░Éņłśņä▒ ĒÆłņóģņ£╝ļĪ£ ļéś ļłäņ¢┤ņ¦äļŗżĻ│Ā ĒĢśņśĆļŗż. ņ┤łņןņØä ļ│┤ļ®┤ ņĀĆļåŹļÅäņØĖ 50mM NaCl ņ▓śļ”¼ĻĄ¼ņØś Ļ▓ĮņÜ░ ņśżļŹ░ņĮöļĪ▒ļ»╝ĒŖĖĻ░Ć ļīĆņĪ░ĻĄ¼ņØś 67% ņłśņżĆņ£╝ļĪ£ Ļ░ÉņåīĒĢśņśĆĻ│Ā, ļŗżļźĖ ļ»╝ĒŖĖļźśļŖö 77~78%ļĪ£ ļ╣äņŖĘĒĢ£ ņłśņżĆņØś ņ┤łņן Ļ░Éņåīņ£©ņØä ļ│┤ņśĆļŗż(Fig. 1). 100mM NaCl ņ▓śļ”¼ĻĄ¼ņŚÉņä£ļŖö ņĢĀĒöīļ»╝ĒŖĖļŖö ļīĆņĪ░ĻĄ¼ņØś 76%ļĪ£, ņŖż Ēö╝ņĢäļ»╝ĒŖĖļŖö 73%, ĒīīņØĖņĢĀĒöīļ»╝ĒŖĖ 72%, ĒÄśĒŹ╝ļ»╝ĒŖĖ 68%, ņśżļŹ░ņĮöļĪ▒ļ»╝ ĒŖĖ 62%, ņ┤łņĮöļ»╝ĒŖĖļŖö 59%ņØś ņłśņżĆņ£╝ļĪ£ ņ┤łņĮöļ»╝ĒŖĖĻ░Ć Ļ░Ćņן ļ¦ÄņØ┤ Ļ░Éņåī ĒĢśņśĆļŗż. ņŚĮ ņāØņןļ¤ēņØä ļ│┤ļ®┤ ņŚĮņןĻ│╝ ņŚĮĒÅŁ ļ¬©ļæÉ ļīĆņĪ░ĻĄ¼ņŚÉ ļ╣äĒĢśņŚ¼ ņŚ╝ņ▓ś ļ”¼ ļåŹļÅäĻ░Ć ņ”ØĻ░ĆĒĢĀņłśļĪØ Ļ░ÉņåīĒĢśņśĆĻ│Ā, ĒÄśĒŹ╝ļ»╝ĒŖĖņÖĆ ņĢĀĒöīļ»╝ĒŖĖļŖö ņĀĆļåŹļÅä ņ▓śļ”¼ņŚÉņä£ļÅä Ļ░ÉņåīĻ░Ć ļÜ£ļĀĘĒĢśņśĆļŗż.
Table┬Ā1.
Effects of NaCl concentration for 3 weeks on the growth of 6 species of mints.
| NaCl (mM) | Plant height (cm) | Leaf length (cm) | Leaf width (cm) |
Fresh weight (g/plant) |
Dry matter (%) |
||
|---|---|---|---|---|---|---|---|
| Shoot | Root | Shoot | Root | ||||
|
|
|||||||
| Peppermint (Mentha ├Ś piperita) | |||||||
| Control | 28.01az | 2.67a | 1.43a | 12.06a | 6.18ab | 12.82b | 9.02a |
| 50 | 21.67b | 1.62b | 0.63b | 11.07b | 5.65b | 16.27a | 8.12a |
| 100 | 19.17b | 1.31b | 0.57b | 11.96ab | 6.77a | 15.78ab | 8.37a |
| 200 | 17.33b | 1.12b | 0.61b | 10.54b | 4.40b | 12.80b | 10.07a |
|
|
|||||||
| Eau de Cologne Mint (Mentha ├Ś piperita f. citrata) | |||||||
| Control | 22.31a | 2.50a | 2.11a | 16.03a | 4.72a | 9.04ab | 8.60a |
| 50 | 14.83b | 2.13ab | 1.60a | 11.99ab | 5.61a | 9.80ab | 6.70b |
| 100 | 13.83b | 1.93ab | 1.53b | 12.32ab | 5.25a | 11.40a | 6.94b |
| 200 | 9.17c | 1.00b | 0.83c | 7.27b | 4.41a | 7.28b | 6.55b |
|
|
|||||||
| Chocomint (Mentha ├Ś piperita f. citrata 'Chocolate') | |||||||
| Control | 18.32a | 2.62a | 1.33a | 12.03b | 4.97b | 11.51c | 8.71b |
| 50 | 14.01b | 2.03b | 1.03b | 15.70a | 6.28ab | 14.80bc | 16.91a |
| 100 | 10.83c | 1.87bc | 0.67c | 11.67b | 6.89ab | 20.25a | 13.46ab |
| 200 | 8.83c | 1.43c | 0.83c | 10.56b | 7.76a | 17.47ab | 10.28ab |
|
|
|||||||
| Spearmint (Mentha spicata) | |||||||
| Control | 17.01a | 2.31a | 1.42a | 10.95ab | 9.11ab | 8.87b | 7.34b |
| 50 | 13.32b | 2.03a | 1.21ab | 12.93a | 11.42a | 9.94b | 8.95ab |
| 100 | 12.38bc | 1.23b | 0.97bc | 12.89a | 8.27b | 13.07a | 9.14a |
| 200 | 11.01c | 0.93b | 0.72c | 8.80b | 8.88b | 12.87b | 8.93ab |
|
|
|||||||
| Applemint (Mentha suaveolens) | |||||||
| Control | 21.83a | 2.12a | 1.47a | 11.81a | 6.75ab | 9.63b | 7.58a |
| 50 | 17.01b | 1.43b | 0.67a | 11.76a | 7.20ab | 11.62b | 7.92a |
| 100 | 16.51b | 1.23b | 0.57a | 10.71a | 9.05a | 14.24a | 6.54a |
| 200 | 12.12c | 1.03b | 0.61a | 8.99b | 4.81b | 10.40b | 5.24a |
|
|
|||||||
| Pineapplemint (Mentha suaveolens 'Variegata') | |||||||
| Control | 20.83a | 3.83a | 1.67a | 9.92a | 5.19a | 11.41b | 10.42a |
| 50 | 16.83b | 2.87b | 1.02ab | 8.66a | 6.31a | 15.76a | 4.71b |
| 100 | 15.02b | 1.97bc | 0.67b | 5.73b | 4.19ab | 13.49ab | 5.28b |
| 200 | 14.53b | 1.31c | 0.61b | 1.95c | 2.19b | 13.46ab | 5.12b |
Fig.┬Ā1.
Effects of NaCl concentration for 3 weeks on the relative plant height of 6 species of mints.
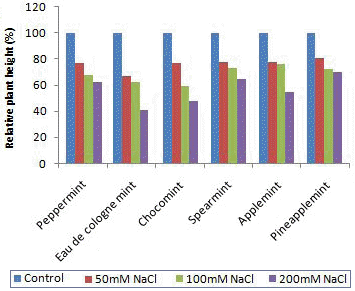
ņ¦ĆņāüļČĆ ņāØņ▓┤ņżæņØś ļ│ĆĒÖöļŖö ņŗżņĀ£ ņØ┤ņÜ® ļČĆņ£äņØĖ ļ¦īĒü╝ ņ×¼ļ░░ ļ░Å ņłśĒÖĢ ņĖĪ ļ®┤ņŚÉņä£ ņżæņÜöĒĢ£ ņÜöņåīņØ┤ļŗż. ņŚ╝ļåŹļÅäņŚÉ ļö░ļźĖ ĒÆłņóģļ│ä ņāØņ▓┤ Ļ░Éņåīņ£©ņØĆ ļŗ¼ ļ×Éņ¦Ćļ¦ī ĒÄśĒŹ╝ļ»╝ĒŖĖ, ņśżļŹ░ņĮöļĪ▒ļ»╝ĒŖĖ, ĒīīņØĖņĢĀĒöīļ»╝ĒŖĖļŖö ļ╣äĻĄÉņĀü ņĀĆļåŹļÅäņŚÉ ņåŹĒĢśļŖö 50mM NaCl ņ▓śļ”¼ĻĄ¼ļČĆĒä░ ņ¦ĆņāüļČĆ ņāØņ▓┤ņżæņØś Ļ░ÉņåīĻ░Ć ņØ╝ņ¢┤ļé¼ ļŗż(Fig. 2). Cuartero and Rafael(1999)ņØĆ ĒåĀļ¦łĒåĀĻ░Ć ņŚ╝ ņŖżĒŖĖļĀłņŖż ļź╝ ļ░øņ£╝ļ®┤ ļ┐īļ”¼ļ│┤ļŗżļŖö ņ¦ĆņāüļČĆņØś ņāØņ£ĪņØ┤ ļŹö ņ¢ĄņĀ£ļÉ£ļŗżĻ│Ā ļ│┤Ļ│ĀĒĢ£ ļ░ö ņ׳ļŗż. ĒĢ£ĒÄĖ ņ┤łņĮöļ»╝ĒŖĖņÖĆ ņŖżĒö╝ņĢäļ»╝ĒŖĖļŖö ļ╣äĻĄÉņĀü ņĀĆļåŹļÅä ņŚ╝ņ▓śļ”¼ĻĄ¼ņŚÉņä£ ļŖö ņśżĒ׳ļĀż ņ¦ĆņāüļČĆ ņāØņ▓┤ņżæņØ┤ ļīĆņĪ░ĻĄ¼ļ│┤ļŗż ņĢĮĻ░ä ņ”ØĻ░ĆĒĢśņśĆĻ│Ā, ĻĘĖņżæ ņŖż Ēö╝ņĢäļ»╝ĒŖĖļŖö 100mM NaCl ņ▓śļ”¼ĻĄ¼ņŚÉņä£ļÅä ļīĆņĪ░ĻĄ¼ļ│┤ļŗż ņ¦ĆņāüļČĆ ņāØņ▓┤ ņżæņØ┤ ņĢĮ 1.18ļ░░ ņ”ØĻ░ĆļÉśņ¢┤ ņŚ╝ņŚÉ ļīĆĒĢ£ ņĀĆĒĢŁņä▒ņØ┤ ņ׳ļŖö Ļ▓āņ£╝ļĪ£ ņāØĻ░üļÉś ņŚłļŗż.
Fig.┬Ā2.
Effects of NaCl concentration for 3 weeks on the relative shoot fresh weight of 6 species of mints.
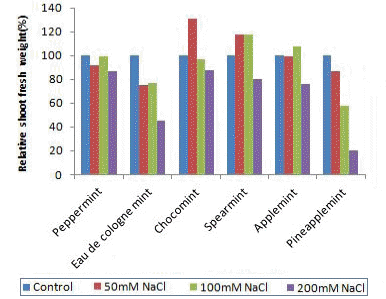
ņ¦ĆĒĢśļČĆ ņāØņ▓┤ņżæņØĆ Ļ│ĀļåŹļÅä ņ▓śļ”¼ĻĄ¼ņØĖ 200mM NaCl ņ▓śļ”¼ĻĄ¼ņŚÉņä£ļŖö Ļ░ÉņåīĒĢśņśĆņ£╝ļéś ņĀĆļåŹļÅä ņ▓śļ”¼ĻĄ¼ņØĖ 50Ļ│╝ 100mM NaCl ņ▓śļ”¼ĻĄ¼ņŚÉņä£ļŖö ņ”ØĻ░ĆĒĢśļŖö Ļ▓ĮĒ¢źņØä ļ│┤ņśĆļŗż. ņØ┤ļ¤¼ĒĢ£ ņĀĆļåŹļÅä ņŚ╝ ņ▓śļ”¼ĻĄ¼ņŚÉņä£ņØś ņ¦ĆĒĢśļČĆ ņāØ ņ▓┤ņżæ ņ”ØĻ░ĆļŖö ĒåĀņ¢æ ļé┤ ņŚ╝ ņ¦æņĀüņ£╝ļĪ£ ņØĖĒĢśņŚ¼ ļåÆņĢäņ¦ä ņé╝Ēł¼ņĢĢ ļĢīļ¼ĖņŚÉ ņøÉĒÖ£ ĒĢśņ¦Ć ļ¬╗ĒĢ£ ļ┐īļ”¼ņØś ņłśļČä ĒØĪņłśļź╝ ĻĘ╣ļ│ĄĒĢśĻĖ░ ņ£äĒĢśņŚ¼ ļ┐īļ”¼ ņāØņ£ĪņØ┤ ņØ╝ņŗ£ņĀü ņ£╝ļĪ£ ņ”ØĻ░ĆĒĢśĻ│Ā ņØ┤ļź╝ ĒåĄĒĢ┤ ņŚ╝ĒĢ┤ļź╝ ņĪ░ņĀłĒĢśļŖö Ļ▓āņ£╝ļĪ£ ņČöņĖĪļÉ£ļŗż. Khan et al.(1999)ņØĆ ĒĢ┤ņĢłĻ░ĆņŚÉ ņä£ņŗØĒĢśļŖö Halopyrum mocoronatum L. ņØś Ļ▓ĮņÜ░ ņé¼Ļ▓Įņ×¼ļ░░ ĒĢśņŚÉņä£ ņŚ╝ ņ▓śļ”¼ļź╝ ĒĢśņśĆņØä Ļ▓ĮņÜ░ ņĀĆļåŹļÅäņØĖ 90mM NaCl ņ▓śļ”¼ĻĄ¼ļŖö ļ┐īļ”¼ņØś ņāØņ▓┤ņżæĻ│╝ Ļ▒┤ļ¼╝ņżæņØ┤ ņśżĒ׳ļĀż ņ”ØĻ░ĆļÉśņŚłĻ│Ā, ĻĘĖ ņØ┤ņāüņØś ļåŹļÅäņŚÉņä£ļŖö Ļ░ÉņåīĒĢśĻĖ░ ņŗ£ņ×æĒĢśņŚ¼ 360mM NaCl ņ▓śļ”¼ņØś Ļ▓ĮņÜ░ ļŖö Ļ│Āņé¼ĒĢśņśĆļŗżĻ│Ā ĒĢśņśĆļŗż.
Ļ▒┤ļ¼╝ļźĀņØä ļ│┤ļ®┤ ņ¦ĆņāüļČĆ ņāØņ▓┤ņżæĻ│╝ļŖö ļŗ¼ļ”¼ ņ¦ĆņāüļČĆ Ļ▒┤ļ¼╝ļźĀņØĆ ņŚ╝ņ▓śļ”¼ ĻĄ¼ņŚÉņä£ ļ¬©ļæÉ ņ”ØĻ░ĆĒĢśļŖö Ļ▓ĮĒ¢źņØ┤ņŚłĻ│Ā, ņ¦ĆĒĢśļČĆņØś Ļ▒┤ļ¼╝ļźĀņØĆ ņŖżĒö╝ņĢäļ»╝ĒŖĖ ņÖĆ ĒŖ╣Ē׳ ņ┤łņĮöļ»╝ĒŖĖļź╝ ņĀ£ņÖĖĒĢśĻ│ĀļŖö ņŚ╝ņ▓śļ”¼ĻĄ¼ņŚÉņä£ ņĀäļ░śņĀüņ£╝ļĪ£ Ļ░ÉņåīĒĢś ļŖö Ļ▓ĮĒ¢źņØä ļ│┤ņśĆļŗż.
ņŚĮļĪØņåī ĒĢ©ļ¤ēņØĆ ņĢĀĒöīļ»╝ĒŖĖ, ņŖżĒö╝ņĢäļ»╝ĒŖĖ, ņ┤łņĮöļ»╝ĒŖĖ ļ░Å ĒÄśĒŹ╝ļ»╝ĒŖĖļŖö ņŚ╝ņ▓śļ”¼ĻĄ¼ņŚÉņä£ ņ”ØĻ░ĆĒĢśņśĆĻ│Ā, ņśżļŹ░ņĮöļĪ▒ļ»╝ĒŖĖņÖĆ ĒīīņØĖņĢĀĒöī ļ»╝ĒŖĖļŖö ņŚ╝ņ▓ś ļ”¼ĻĄ¼ņŚÉņä£ Ļ░ÉņåīĒĢśļŖö Ļ▓ĮĒ¢źņØä ļ│┤ņśĆļŗż(Fig. 3). ņŚĮļĪØņåī ņ”ØĻ░ĆĒśäņāüņØĆ ņØ╝ ļ░śņĀüņ£╝ļĪ£ ņŚ╝ ņŖżĒŖĖļĀłņŖż ĒĢśņŚÉņä£ ņŚĮļĪØņåī ĒĢ©ļ¤ēņØ┤ ļ¢©ņ¢┤ņ¦äļŗżļŖö ļ│┤Ļ│ĀņÖĆļŖö ņāüļ░śļÉ£ Ļ▓░Ļ│╝ņśĆņ£╝ļéś(Parida and Das, 2005), Kang and Shim(1998) ņØ┤ ļ│┤Ļ│ĀĒĢ£ Ļ▓āĻ│╝ Ļ░ÖņØ┤ ņŗ¼ĒĢśĻ▓ī ņāØņ£ĪņØ┤ ņ¢ĄņĀ£ļÉśņ¢┤ ņ×ÄņØś ļģĖĒÖö ļ░Å Ļ│Āņé¼Ļ░Ć ņØ╝ņ¢┤ļéśĻĖ░ ņĀäņØś ņ┤łĻĖ░ ņŚ╝ ņ▓śļ”¼ ņāüĒā£ņŚÉņä£ļŖö ņŚĮļé┤ ņŚĮļĪØņåīņØś ĒĢ©ļ¤ēņØ┤ ņØ╝ ņŗ£ņĀüņ£╝ļĪ£ ņ”ØĻ░ĆļÉ£ļŗżļŖö ļ│┤Ļ│ĀņÖĆ ņ£Āņé¼ĒĢ£ Ļ▓░Ļ│╝ļĪ£ ņāØĻ░üļÉ£ļŗż.
Fig.┬Ā3.
Effects of NaCl concentration on the chlorophyll content measured by SPAD of peppermint (Mentha ├Ś piperita)(A), eau de cologne mint (Mentha ├Ś piperita f. citrata)(B), chocomint (Mentha ├Ś piperita f. citrata 'Chocolate')(C), spearmint (Mentha spicata)(D), applemint (Mentha suaveolens)(E) and pineapplemint (Mentha suaveolens 'Variegata')(F).
zVertical bars represent ┬▒ SD of the means (n=10).
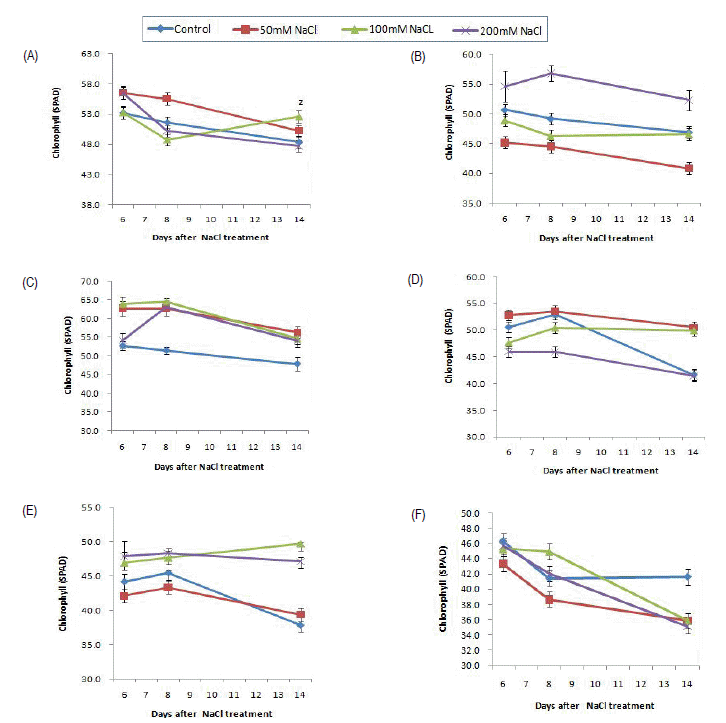
ĒŖ╣Ē׳ ņ¦äĒĢ£ ņĢöņ×ÉņāēņØä ļØäļŖö ņ┤łņĮöļ»╝ĒŖĖļŖö ļīĆņĪ░ĻĄ¼ņØś 47.28ņŚÉ ļ╣äĒĢśņŚ¼ 50mM NaCl ņ▓śļ”¼ĻĄ¼ 56.27, 100mM NaCl ņ▓śļ”¼ĻĄ¼ 54.59, 200mM NaCl ņ▓śļ”¼ĻĄ¼ 54.07ļĪ£ ļŹö ļåÆĻ▓ī ļéśĒāĆļé¼ļŗż. Kennedy and De-Fillippis(1999)ļŖö GrevileaņŚÉņä£ ņŚ╝ ņŖżĒŖĖļĀłņŖż ņ▓śļ”¼ņŚÉ ņØśĒĢ┤ ņŚĮļĪØņåī ņÖĆ ņ╣┤ļĪ£Ēŗ░ļģĖņØ┤ļō£ ņĀäņ▓┤ ĒĢ©ļ¤ēņØĆ ņżäņ¢┤ļōżņŚłņ¦Ćļ¦ī ņĢłĒåĀņŗ£ņĢäļŗīņØś ĒĢ©ļ¤ēņØĆ ņ£ĀņØśņĀüņ£╝ļĪ£ ņ”ØĻ░ĆĒĢ£ļŗżĻ│Ā ĒĢśņśĆļŖöļŹ░, ņ┤łņĮöļ»╝ĒŖĖņØś Ļ▓ĮņÜ░ ņŚ╝ ņ▓śļ”¼ņŚÉ ņØśĒĢ£ ņĢłĒåĀņŗ£ņĢäļŗī ĒĢ©ļ¤ē ņ”ØĻ░ĆĻ░Ć ņśłņāüļÉśļ®░ ņØ┤ņŚÉ ļīĆĒĢ£ ņČöĻ░Ć ņŚ░ĻĄ¼Ļ░Ć ĒĢäņÜöĒĢśļŗż Ļ│Ā ļ│Ėļŗż. ĒĢ£ĒÄĖ ņŚ╝ ņČĢņĀüņØĆ ņןĻĖ░ņĀüņ£╝ļĪ£ļŖö ņŚĮļĪØņåīņØś Ļ░Éņåī(Santos, 2004), Ļ┤æĒĢ®ņä▒ ĒÜ©ņ£©ņØś Ļ░Éņåī(Munns, 2002) ļō▒ņØä ņ£Āļ░£ĒĢśļŖöļŹ░, ņśżļŹ░ ņĮöļĪ▒ ļ»╝ĒŖĖņÖĆ ĒīīņØĖņĢĀĒöī ļ»╝ĒŖĖņØś Ļ▓ĮņÜ░ ņŚĮļĪØņåīĻ░Ć Ļ░ÉņåīĒĢśņśĆļŗż. ĒŖ╣Ē׳ ņŚ░ ļģĖļ×æ~ņŚ░ļģ╣ņāē ļ░śņŚĮņóģņØĖ ĒīīņØĖņĢĀĒöīļ»╝ĒŖĖņØś ņŚĮ ĒŖ╣ņä▒ņØä Ļ│ĀļĀżĒĢĀ ļĢī ĒśäņĀĆ ĒĢ£ ņŚĮļĪØņåī Ļ░ÉņåīļĪ£ ņØĖĒĢ£ ņāØļ”¼ņĀü ļīĆņé¼ ļ│ĆĒÖöĻ░Ć ņśłņāüļÉśņŚłļŗż. ņØ╝ļ░śņĀüņ£╝ ļĪ£ ņŗØļ¼╝ņØĆ ņŚ╝ ņŖżĒŖĖļĀłņŖżļź╝ ļ░øņ£╝ļ®┤ ĻĖ░Ļ│ĄņØ┤ ļŗ½Ē׳Ļ│Ā Ļ┤æĒĢ®ņä▒ ĒÜ©ņ£©ņØ┤ ļ¢© ņ¢┤ņ¦ĆļŖöļŹ░(Parida et al., 2002), Chon and Park(2003)ņØĆ ņŚ╝ļåŹļÅäĻ░Ć ņ”ØĻ░ĆĒĢĀņłśļĪØ ņĮ®ņØś ņŚĮļĪØņåī ĒĢ©ļ¤ēĻ│╝ Ļ┤æĒĢ®ņä▒ ĒÜ©ņ£©ņØ┤ ļ¢©ņ¢┤ņ¦äļŗżĻ│Ā ĒĢśņśĆĻ│Ā, Choi et al.(1997)ļÅä ņŚ╝ļåŹļÅäĻ░Ć ņ”ØĻ░ĆĒĢĀņłśļĪØ ļ│┤ļ”¼ņØś ņŚĮļĪØņåī ĒĢ©ļ¤ēņØ┤ ļ¢©ņ¢┤ņ¦äļŗżĻ│Ā ļ░ØĒśöļŗż.
ĒöīļØ╝ļ│┤ļģĖņØ┤ļō£ļź╝ ĒżĒĢ©ĒĢ£ ņŗØļ¼╝ņ▓┤ ļé┤ņØś ĒÄśļåĆĒÖöĒĢ®ļ¼╝ņØĆ ņŚ¼ļ¤¼ Ļ░Ćņ¦Ć ņŖż ĒŖĖļĀłņŖż ņÜöņØĖņŚÉ ļ░śņØæĒĢśņŚ¼ ņŗØļ¼╝ņØ┤ ĒĢŁņāüņä▒ ņ£Āņ¦Ćļź╝ ņ£äĒĢ┤ ņāØņä▒ĒĢśļŖö 2ņ░© ļīĆņé¼ņé░ļ¼╝ļĪ£ņä£ ĒĢŁņé░ĒÖö, ĒĢŁĻĘĀ ļō▒ ļŗżņ¢æĒĢ£ ņāØļ”¼ņĀü ļ░śņØæņØä ļéśĒāĆļéĖļŗż (Madsen and Bertelsen, 1995; Cao and Cao, 1999; Kivilomopolo and Hyotylainen, 2007). ņŚ¼ļ¤¼ Ļ░Ćņ¦Ć ņŖżĒŖĖļĀłņŖż ņÜöņØĖ ņżæņØś ĒĢśļéśņØĖ ņŚ╝ ņŖżĒŖĖļĀłņŖżļŖö ņé░ĒÖöņĀü ņŖżĒŖĖļĀłņŖż ļśÉĒĢ£ ņ£Āļ░£ņŗ£ĒéżļŖöļŹ░, ņŗØļ¼╝ņØĆ ņØ┤ļĪ£ ņØĖĒĢś ņŚ¼ Ļ│╝ļŗżĒĢśĻ▓ī ļ░£ņāØļÉśļŖö ĒÖ£ņä▒ ņé░ņåīņóģņØä ņĀ£Ļ▒░ĒĢśĻĖ░ ņ£äĒĢ┤ ĒĢŁņé░ĒÖö ĒÜ©ņåīļ┐É ļ¦ī ņĢäļŗłļØ╝ 2ņ░©ļīĆņé¼ņé░ļ¼╝ņØĖ ĒĢŁņé░ĒÖöļ¼╝ņ¦ł ļśÉĒĢ£ ņāØņä▒, ņČĢņĀüņŗ£Ēé©ļŗż (Alsher and Hess, 1993; Last et al., 1997). ļ│Ė ņŗżĒŚś Ļ▓░Ļ│╝ņŚÉņä£ļÅä ĒÆł ņóģļ│ä, ņŚ╝ ļåŹļÅäļ│äļĪ£ ņ”ØĻ░Ćļ¤ēņØĆ ņ░©ņØ┤Ļ░Ć ņ׳ņŚłņ¦Ćļ¦ī ļīĆņ▓┤ļĪ£ ņŚ╝ ņ▓śļ”¼ĻĄ¼ļōż ņØ┤ ļīĆņĪ░ĻĄ¼ņŚÉ ļ╣äĒĢ┤ ĒĢŁņé░ĒÖö ĒÖ£ņä▒ ļ░Å ņ┤Ø ĒÅ┤ļ”¼ĒÄśļåĆ, ņ┤Ø ĒöīļØ╝ļ│┤ļģĖņØ┤ļō£ņØś ĒĢ©ļ¤ēņØ┤ ņ”ØĻ░ĆĒĢ£ Ļ▓āņØä ĒÖĢņØĖĒĢĀ ņłś ņ׳ņŚłļŗż(Table 2).
Table┬Ā2.
Effects of NaCl concentration for 3 weeks on the DPPH (1,1- diphenyl-2-picrylhydrazyl) radical scavenging activity, total phenolics, and total flavonoids of 6 species of mints.
| NaCl (mM) | DPPH assayz (EDA%)y | Total phenolics (┬ĄgŌĆómL-1) | Total flavonoids (mgŌĆómL-1) |
|---|---|---|---|
|
|
|||
| Peppermint (Mentha ├Ś piperita) | |||
| Control | 25.73cx | 12.79b | 2.86b |
| 50 | 35.16b | 17.88a | 1.24d |
| 100 | 41.46a | 26.50a | 2.66c |
| 200 | 32.89b | 21.58a | 4.09a |
|
|
|||
| Eau de Cologne Mint (Mentha ├Ś piperita f. citrata) | |||
| Control | 28.45c | 12.71c | 1.27c |
| 50 | 51.62b | 32.69a | 2.92a |
| 100 | 71.24a | 30.44a | 2.39b |
| 200 | 46.45b | 24.67b | 3.06a |
|
|
|||
| Chocomint (Mentha ├Ś piperita f. citrata 'Chocolate') | |||
| Control | 84.14b | 30.52b | 4.59b |
| 50 | 87.03a | 81.27a | 5.09a |
| 100 | 87.83a | 88.98a | 5.07a |
| 200 | 88.03a | 85.08a | 5.08a |
|
|
|||
| Spearmint (Mentha spicata) | |||
| Control | 41.48c | 76.04a | 4.94a |
| 50 | 85.34a | 40.48b | 5.09a |
| 100 | 47.06c | 67.79a | 4.62a |
| 200 | 69.86b | 41.56b | 1.75b |
|
|
|||
| Applemint (Mentha suaveolens) | |||
| Control | 32.45a | 18.69a | 0.92c |
| 50 | 34.98a | 12.75b | 0.91c |
| 100 | 38.92a | 7.23c | 1.67b |
| 200 | 30.60a | 14.83b | 2.41a |
|
|
|||
| Pineapplemint (Mentha suaveolens 'Variegata') | |||
| Control | 63.61bc | 24.98b | 3.56a |
| 50 | 64.89b | 24.90b | 2.93b |
| 100 | 59.47c | 24.04b | 2.37b |
| 200 | 82.92a | 43.83a | 2.35c |
ļ»╝ĒŖĖ ĒÆłņóģļ│ä ĒĢŁņé░ĒÖö ĒÖ£ņä▒ņØä DPPH ļØ╝ļööņ╣╝ ņåīĻ▒░ļ▓ĢņØä ņØ┤ņÜ®ĒĢśņŚ¼ ņĖĪ ņĀĢĒĢ£ Ļ▓░Ļ│╝ ĒĢŁņé░ĒÖö ĒÖ£ņä▒ņØ┤ Ļ░Ćņן ļåÆņĢśļŹś ĒÆłņóģņØĆ ņ┤łņĮöļ»╝ĒŖĖņśĆļŗż. ņ┤łņĮö ļ»╝ĒŖĖļŖö ļīĆņĪ░ĻĄ¼ 84.14%, 50mM NaCl ņ▓śļ”¼ĻĄ¼ 87.03%, 100mM NaCl ņ▓śļ”¼ĻĄ¼ 87.83%, 200mM NaCl ņ▓śļ”¼ĻĄ¼ 88.03% ņł£ņ£╝ļĪ£ ņŚ╝ ņ▓ś ļ”¼ ņŗ£ ĒĢŁņé░ĒÖö ĒÖ£ņä▒ņØ┤ ņ”ØĻ░ĆĒĢśļŖö Ļ▓ĮĒ¢źņØ┤ņŚłņ£╝ļéś ņØ┤ļ¤¼ĒĢ£ ņ┤łņĮöļ»╝ĒŖĖņØś ļåÆ ņØĆ ĒĢŁņé░ĒÖö ĒÖ£ņä▒ņØĆ ņŚ╝ ņ▓śļ”¼ņŚÉ ļö░ļźĖ ņé░ĒÖöņĀü ņŖżĒŖĖļĀłņŖżļź╝ ņ¢ĄņĀ£ĒĢśĻĖ░ ņ£ä ĒĢ£ Ļ▓āļ│┤ļŗżļŖö ņ┤łņĮöļ»╝ĒŖĖ ĒÆłņóģ ņ×Éņ▓┤ņØś ĒĢŁņé░ĒÖö ĒÖ£ņä▒ņØ┤ ļåÆņØĆ ĒŖ╣ņä▒ņŚÉ ĻĖ░ ņØĖĒĢ£ Ļ▓āņ£╝ļĪ£ ņāØĻ░üļÉ£ļŗż. ņŚ╝ņ▓śļ”¼ņŚÉ ņØśĒĢ┤ ĒĢŁņé░ĒÖö ĒÖ£ņä▒ ļ│ĆĒÖöĻ░Ć Ļ░Ćņן ļÜ£ ļĀĘĒĢśņśĆļŹś ņŖżĒö╝ņĢäļ»╝ĒŖĖļŖö ĒŖ╣Ē׳ 50mM NaCl ņ▓śļ”¼ĻĄ¼ņŚÉņä£ 85.34%ļĪ£ ĒĢŁņé░ĒÖö ĒÖ£ņä▒ņØ┤ Ļ░Ćņן ļåÆņĢśĻ│Ā 200mM ņ▓śļ”¼ĻĄ¼ņŚÉņä£ļÅä ļåÆņØĆ ĒÖ£ņä▒ņØä ļ│┤ ņśĆļŗż. ĒīīņØĖņĢĀĒöīļ»╝ĒŖĖņØś ĒĢŁņé░ĒÖö ĒÖ£ņä▒ļÅä 100mM NaCl ņ▓śļ”¼ĻĄ¼Ļ╣īņ¦ĆļŖö ļīĆņĪ░ĻĄ¼ 63.61%ņÖĆņØś ņ░©ņØ┤Ļ░Ć Ļ▒░ņØś ņŚåņŚłņ£╝ļéś 200mM NaCl ņ▓śļ”¼ĻĄ¼ ņŚÉņä£ ĒĢŁņé░ĒÖö ĒÖ£ņä▒ņØ┤ 82.92%ļĪ£ ņ”ØĻ░ĆļÉśļŖö Ļ▓āņ£╝ļĪ£ ļéśĒāĆļé¼ļŗż. ņśżļŹ░ņĮö ļĪ▒ļ»╝ĒŖĖ, ĒÄśĒŹ╝ļ»╝ĒŖĖ, ņĢĀĒöīļ»╝ĒŖĖļŖö 100mM NaCl ņ▓śļ”¼ĻĄ¼Ļ╣īņ¦ĆļŖö ņŚ╝ļåŹ ļÅäĻ░Ć ļåÆņĢäņ¦łņłśļĪØ ĒĢŁņé░ĒÖö ĒÖ£ņä▒ņØ┤ ņ”ØĻ░ĆļÉśņŚłĻ│Ā Ļ│ĀļåŹļÅäņ▓śļ”¼ĻĄ¼ņØĖ 200mM NaCl ņ▓śļ”¼ĻĄ¼ņŚÉņä£ ļŗżņŗ£ ĒĢŁņé░ĒÖö ĒÖ£ņä▒ņØ┤ Ļ░ÉņåīĒĢśļŖö Ļ▓ĮĒ¢źņØ┤ ļéś ĒāĆļé¼ņ£╝ļ®░, ņĢĀĒöīļ»╝ĒŖĖņØś ĒĢŁņé░ĒÖö ĒÖ£ņä▒ņØĆ ļŗżļźĖ ĒÆłņóģņŚÉ ļ╣äĒĢ┤ ļé«ņĢśļŗż.
ņŚ╝ļåŹļÅäļ│ä ņ┤Ø ĒÅ┤ļ”¼ĒÄśļåĆņØś ĒĢ©ļ¤ē ļ│ĆĒÖöļź╝ ļ╣äĻĄÉĒĢ£ Ļ▓░Ļ│╝ ņĀäļ░śņĀüņ£╝ļĪ£ ņ┤Ø ĒÄśļåĆ ĒÖöĒĢ®ļ¼╝ ĒĢ©ļ¤ēņØ┤ ļåÆņĢśļŹś ļ»╝ĒŖĖļźśļŖö ņ┤łņĮöļ»╝ĒŖĖņÖĆ ņŖżĒö╝ņĢäļ»╝ĒŖĖļĪ£, ņØ┤ļŖö ĒĢŁņé░ĒÖö ĒÖ£ņä▒ ļśÉĒĢ£ ļåÆņĢśļŹś ĒÆłņóģņØ┤ņŚłļŗż. ļśÉĒĢ£ ņśżļŹ░ņĮöļĪ▒ļ»╝ĒŖĖņØś ņ┤Ø ĒÅ┤ļ”¼ĒÄśļåĆĒÖöĒĢ®ļ¼╝ ĒĢ©ļ¤ēņØĆ ņŚ╝ ņ▓śļ”¼ĻĄ¼Ļ░Ć ļīĆņĪ░ĻĄ¼ļ│┤ļŗż ļåÆņĢśļŖöļŹ░, ĻĘĖņżæ 50mM NaCl ņ▓śļ”¼ĻĄ¼Ļ░Ć Ļ░Ćņן ļåÆņØĆ ĒĢ©ļ¤ēņØä ļéśĒāĆļé┤ņŚłĻ│Ā ĻĘĖ ņØ┤ņāüņØś ņŚ╝ ļåŹļÅäņŚÉņä£ļŖö ņĀÉņ░© Ļ░ÉņåīĒĢśļŖö Ļ▓āņ£╝ļĪ£ ļéśĒāĆļé¼ļŗż. ĒīīņØĖņĢĀĒöīļ»╝ĒŖĖņØś Ļ▓ĮņÜ░ ļŖö 100mM NaCl ņ▓śļ”¼ĻĄ¼Ļ╣īņ¦ĆļŖö ļīĆņĪ░ĻĄ¼ņÖĆ ņ£Āņé¼ĒĢśņśĆņ£╝ļéś 200mM NaCl ņ▓śļ”¼ĻĄ¼ņŚÉņä£ ļīĆņĪ░ĻĄ¼ņØś 1.75 ļ░░ļĪ£ ņ”ØĻ░ĆĒĢśņśĆļŗż. ĒÄśĒŹ╝ļ»╝ĒŖĖņÖĆ ņĢĀ Ēöīļ»╝ĒŖĖņØś ņ┤Ø ĒÅ┤ļ”¼ĒÄśļåĆ ĒÖöĒĢ®ļ¼╝ ĒĢ©ļ¤ēņØĆ ļŗżļźĖ ļ»╝ĒŖĖļźśņŚÉ ļ╣äĒĢśņŚ¼ ļé«ņØĆ ĒÄĖņŚÉ ņåŹĒĢśņśĆļŖöļŹ░, ĻĘĖņżæņŚÉņä£ļÅä ņĢĀĒöīļ»╝ĒŖĖņØś ņ┤Ø ĒÅ┤ļ”¼ĒÄśļåĆĒÖöĒĢ®ļ¼╝ ĒĢ©ļ¤ē ņØ┤ ļŹö ļé«ņĢśņ£╝ļ®░, ĒÄśĒŹ╝ļ»╝ĒŖĖļŖö 100mM NaCl ņ▓śļ”¼ĻĄ¼Ļ╣īņ¦ĆļŖö ņŚ╝ ņ▓śļ”¼ ļåŹļÅäĻ░Ć ļåÆņĢäņ¦ÉņŚÉ ļö░ļØ╝ ļīĆņĪ░ĻĄ¼ņŚÉ ļ╣äĒĢ┤ ņ┤Ø ĒÅ┤ļ”¼ĒÄśļåĆĒÖöĒĢ®ļ¼╝ ĒĢ©ļ¤ēņØ┤ ņ”Ø Ļ░ĆĒĢśņśĆļŗż. ņŚ╝ ņ▓śļ”¼ņŚÉ ļö░ļźĖ ĒĢŁņé░ĒÖö ĒÖ£ņä▒Ļ│╝ ņ┤Ø ĒÅ┤ļ”¼ĒÄśļåĆ ĒÖöĒĢ®ļ¼╝ ĒĢ©ļ¤ē ļ│ĆĒÖöļź╝ ņŚ░Ļ┤Ćņŗ£ņ╝£ Ļ│Āņ░░ĒĢ┤ļ│┤ļ®┤ ļīĆļČĆļČä ņŚ╝ ņ▓śļ”¼ ņŗ£ ļīĆņĪ░ĻĄ¼ņŚÉ ļ╣äĒĢ┤ ĒĢŁ ņé░ĒÖö ĒÖ£ņä▒ņØ┤ ņ”ØĻ░ĆĒĢśņśĆĻ│Ā, ņ┤Ø ĒÅ┤ļ”¼ĒÄśļåĆĒÖöĒĢ®ļ¼╝ ĒĢ©ļ¤ē ļśÉĒĢ£ ņ”ØĻ░ĆĒĢśļŖö Ļ▓ĮĒ¢źņØ┤ ļéśĒāĆļé¼ļŖöļŹ░, ņŚ¼ļ¤¼ ņŚ░ĻĄ¼Ļ▓░Ļ│╝ņŚÉņä£ļÅä ĒĢŁņé░ĒÖö ĒÖ£ņä▒ ļ░Å ņ┤Ø ĒÅ┤ļ”¼ ĒÄśļåĆ ĒÖöĒĢ®ļ¼╝ ĒĢ©ļ¤ēņØś ņāüĻ┤Ćņä▒ņØ┤ ļ│┤Ļ│ĀļÉ£ ļ░ö ņ׳ļŗż(Choi, 2001; Valko et al., 2007; Park, 2008; Woo et al., 2010). ĒĢ£ĒÄĖ Park (2008)ņØĆ Ļ┐ĆĒÆĆĻ│╝ņØś ļ░öņ¦ł(Ocimum basilicum)ņØĆ ĒÆłņóģĻ░ä ņ┤Ø ĒÅ┤ļ”¼ĒÄśļåĆ ĒĢ©ļ¤ēņŚÉ ņāüļŗ╣ĒĢ£ ņ░©ņØ┤Ļ░Ć ņ׳ļŗżĻ│Ā ĒĢśņśĆļŗż.
Lee et al.(2001)ņØĆ ļ»╝ĒŖĖļźśĻ░Ć ļ╣äĻĄÉņĀü ļåÆņØĆ ĒöīļØ╝ļ│┤ļģĖņØ┤ļō£ ĒĢ©ļ¤ēņØä Ļ░Ćņ¦ĆĻ│Ā ņ׳ļŗżĻ│Ā ļ│┤Ļ│ĀĒĢśņśĆļŖöļŹ░, ļ»╝ĒŖĖļźśņØś ņ┤Ø ĒöīļØ╝ļ│┤ļģĖņØ┤ļō£ ĒĢ©ļ¤ē ļ│ĆĒÖö ļź╝ ļ│┤ļ®┤ ĒīīņØĖņĢĀĒöīļ»╝ĒŖĖļź╝ ņĀ£ņÖĖĒĢśĻ│ĀļŖö ļīĆņ▓┤ņĀüņ£╝ļĪ£ ņŚ╝ ņ▓śļ”¼ ņŗ£ ņ┤Ø Ēöī ļØ╝ļ│┤ļģĖņØ┤ļō£ņØś ĒĢ©ļ¤ēņØ┤ ņ”ØĻ░ĆĒĢśļŖö Ļ▓āņ£╝ļĪ£ ļéśĒāĆļé¼ļŗż. ņ┤Ø ĒöīļØ╝ļ│┤ļģĖņØ┤ļō£ ĒĢ©ļ¤ēņØ┤ ļåÆņØĆ ĒÄĖņŚÉ ņåŹĒĢśņśĆļŹś ĒÆłņóģņØĆ ņ┤łņĮöļ»╝ĒŖĖņÖĆ ņŖżĒö╝ņĢäļ»╝ĒŖĖļź╝ ļōż ņłś ņ׳ņŚłļŖöļŹ░, ņ┤łņĮöļ»╝ĒŖĖļŖö ņŚ╝ņ▓śļ”¼ĻĄ¼ ļōżņØ┤ ļīĆņĪ░ĻĄ¼ļ│┤ļŗż ņ┤Ø ĒöīļØ╝ļ│┤ļģĖņØ┤ļō£ ņØś ĒĢ©ļ¤ēņØĆ ņ”ØĻ░ĆļÉśņŚłņ¦Ćļ¦ī ņŚ╝ ļåŹļÅäļ│ä ņ░©ņØ┤ļŖö ļéśĒāĆļéśņ¦Ć ņĢŖņĢśĻ│Ā, ņŖżĒö╝ ņĢäļ»╝ĒŖĖļŖö 50mM NaCl ņ▓śļ”¼ĻĄ¼ņØś ņ┤Ø ĒöīļØ╝ļ│┤ļģĖņØ┤ļō£ņØś ĒĢ©ļ¤ēņØ┤ 5.09mgŌłÖmL-1ļĪ£ņä£ ļīĆņĪ░ĻĄ¼ 4.94mgŌłÖmL-1ļ│┤ļŗż ņåīĒÅŁ Ļ░ÉņåīĒĢśņśĆņ¦Ćļ¦ī ĻĘĖ ņØ┤ņāüņØś ņŚ╝ ļåŹļÅä ņ▓śļ”¼ĻĄ¼ņŚÉņä£ļŖö ļīĆņĪ░ĻĄ¼ļ│┤ļŗż ļé«ņĢäņ¦ĆļŖö Ļ▓ĮĒ¢źņØä ļéśĒāĆ ļé┤ņŚłļŗż. ļśÉĒĢ£ ĒīīņØĖņĢĀĒöīļ»╝ĒŖĖļŖö ļīĆņĪ░ĻĄ¼ņØś ņ┤Ø ĒöīļØ╝ļ│┤ļģĖņØ┤ļō£ ĒĢ©ļ¤ēņØ┤ Ļ░Ć ņן ļåÆņĢśĻ│Ā ņŚ╝ ņ▓śļ”¼ ļåŹļÅäĻ░Ć ļåÆņĢäņ¦ÉņŚÉ ļö░ļØ╝ ĒöīļØ╝ļ│┤ļģĖņØ┤ļō£ņØś ĒĢ©ļ¤ēņØ┤ ņżäņ¢┤ļō£ļŖö Ļ▓āņ£╝ļĪ£ ļéśĒāĆļé¼ņ£╝ļéś ņśżļŹ░ņĮöļĪ▒ļ»╝ĒŖĖ, ĒÄśĒŹ╝ļ»╝ĒŖĖ, ņĢĀĒöīļ»╝ĒŖĖņØś Ļ▓ĮņÜ░ļŖö ņØ┤ņÖĆ ļ░śļīĆļĪ£ ņŚ╝ ļåŹļÅäĻ░Ć ļåÆņØĆ ņ▓śļ”¼ĻĄ¼ņØ╝ņłśļĪØ ļīĆņĪ░ĻĄ¼ ĒöīļØ╝ļ│┤ļģĖ ņØ┤ļō£ ĒĢ©ļ¤ēļ│┤ļŗż ļåÆņĢäņ¦ĆļŖö Ļ▓ĮĒ¢źņØä ļéśĒāĆļāłļŗż.
ņØ┤ņāüņØś ņŗżĒŚśĻ▓░Ļ│╝ļź╝ ņóģĒĢ®ĒĢ┤ļ│┤ļ®┤ ņŚ╝ ļåŹļÅä ņ▓śļ”¼ņŚÉ ļö░ļźĖ ļ»╝ĒŖĖļźśņØś ņāØņ£ĪņØĆ ņŚ╝ ļåŹļÅäĻ░Ć ļåÆņĢäņ¦łņłśļĪØ ņ¢ĄņĀ£ļÉ£ ļ░śļ®┤, ņ¦ĆņāüļČĆ ņāØņ▓┤ņżæņØĆ ņĀĆļåŹ ļÅäņŚÉņä£ļŖö ļīĆņĪ░ĻĄ¼ļ│┤ļŗż ņĢĮĻ░ä ņ”ØĻ░ĆļÉśļŖö Ļ▓ĮĒ¢źņØ┤ ļéśĒāĆļé¼Ļ│Ā, ņ¦ĆņāüļČĆ Ļ▒┤ļ¼╝ ņżæņØĆ ņŚ╝ņ▓śļ”¼ĻĄ¼ņŚÉņä£ ļÜ£ļĀĘĒĢśĻ▓ī ņ”ØĻ░ĆĒĢśņśĆļŗż. ĒĢŁņé░ĒÖö ĒÖ£ņä▒ ļ░Å ņ┤Ø ĒÅ┤ļ”¼ ĒÄśļåĆņØś ĒĢ©ļ¤ēļÅä ņŚŁņŗ£ ņ”ØĻ░ĆĒĢśļŖö Ļ▓ĮĒ¢źņØ┤ ļéśĒāĆļé¼ļŗż. ļśÉĒĢ£ ņ┤Ø ĒöīļØ╝ļ│┤ļģĖ ņØ┤ļō£ ĒĢ©ļ¤ēņØĆ ĒÆłņóģņŚÉ ļö░ļØ╝ ņŚ╝ņ▓śļ”¼ņŚÉ ņØśĒĢ┤ ļīĆņĪ░ĻĄ¼ļ│┤ļŗż ĒĢ©ļ¤ēņØ┤ ņ”ØĻ░ĆļÉś ĻĖ░ļÅä ĒĢśĻ│Ā ņśżĒ׳ļĀż Ļ░ÉņåīļÉśĻĖ░ļÅä ĒĢśņśĆļŗż. Kim et al.(2010)ņØĆ ļ░░ņČöņØś ņŚ╝ ņŖżĒŖĖļĀłņŖż ņ▓śļ”¼ ņŗ£ ņĀĆļåŹļÅäņØĖ 50mM NaCl ņ▓śļ”¼ļŖö ņśżĒ׳ļĀż ņāØņןņØä ņ┤ēņ¦äĒĢśĻ│Ā ĻĖĆļŻ©ĒāĆņ╣śņś© ļ░Å ļ╣äĒāĆļ»╝ C ĒĢ©ļ¤ēņØä ņ”ØĻ░Ćņŗ£Ēéżļ»ĆļĪ£ ņŗØļ¼╝ņØś ņāØ ņן ņ┤ēņ¦äņŚÉ ņØśĒĢ£ ņ×¼ļ░░ ĻĖ░Ļ░äņØś ļŗ©ņČĢ ĒÜ©Ļ│╝ļÅä ĻĖ░ļīĆļÉ£ļŗżĻ│Ā ĒĢśņśĆļŖöļŹ░, ļ│Ė ņŗżĒŚś Ļ▓░Ļ│╝ņŚÉņä£ļÅä ņĀĆļåŹļÅä ņŚ╝ ņŖżĒŖĖļĀłņŖż ņ▓śļ”¼ņŚÉņä£ļŖö ņśżĒ׳ļĀż ĒĢŁņé░ĒÖö ĒÖ£ ņä▒, ņ┤Ø ĒÅ┤ļ”¼ĒÄśļåĆ ĒĢ©ļ¤ē, ņ┤Ø ĒöīļØ╝ļ│┤ļģĖņØ┤ļō£ ĒĢ©ļ¤ēņØ┤ ļīĆņĪ░ĻĄ¼ļ│┤ļŗż ņ”ØĻ░ĆļÉś ļŖö Ļ▓ĮĒ¢źņØä ļéśĒāĆļé┤ņŚłļŗż. ņØ┤ņ▓śļ¤╝ ņĀüņĀłĒĢ£ ņŚ╝ ņ▓śļ”¼ļŖö ņŗØļ¼╝ņØś ļ░®ņ¢┤ĻĖ░ņ×æ ņØä ņ×ÉĻĘ╣ņŗ£ņ╝£ 2ņ░© ļīĆņé¼ņé░ļ¼╝ņØĖ ĒĢŁņé░ĒÖö ņä▒ļČäņØä ņ”Øņ¦äņŗ£ĒéżĻ│Ā ņĢäņÜĖļ¤¼ ņāØ ņ£Ī ņ┤ēņ¦äĒÜ©Ļ│╝Ļ░Ć ļ░£ņāØĒĢśĻ▓ī ļÉśļ»ĆļĪ£ ĒŚłļĖīņŗØļ¼╝ņØś ņ×¼ļ░░ ņŗ£ ĒÖ£ņÜ®Ļ░Ćņ╣śĻ░Ć ļåÆ ņØĆ Ļ▓ĮņóģĻĖ░ņłĀ ņÜöņåīĻ░Ć ļÉĀ ņłś ņ׳ņØä Ļ▓āņØ┤ļŗż. ĒŖ╣Ē׳ ĒĢŁņé░ĒÖöņä▒ņØ┤ ņØ┤ļ»Ė ļ│┤Ļ│Ā ļÉ£ ļ»╝ĒŖĖļźśņØś Ļ▓ĮņÜ░ ļŹöņÜ▒ ņäĖļČäĒÖöļÉ£ ņŗżĒŚśņØä ĒåĄĒĢ┤ Ļ░äņ▓Öņ¦Ćļéś ņŚ╝ ņČĢņĀüņ£╝ ļĪ£ ņØĖĒĢśņŚ¼ ņ×æļ¼╝ ņ×¼ļ░░Ļ░Ć ņ¢┤ļĀżņÜ┤ ņ¦ĆņŚŁņŚÉņä£ Ļ░äņ×æņŗØļ¼╝ļĪ£ļÅä ĒÖ£ņÜ®ņØ┤ Ļ░ĆļŖź ĒĢśļ®░ ņĢäņÜĖļ¤¼ Ē¢źņØä ĒÖ£ņÜ®ĒĢ£ Ļ│ĀĻĖ░ļŖźņä▒ ĒŚłļĖī ņ×¼ļ░░ļŗ©ņ¦Ć ņĪ░ņä▒ņØä ņ£äĒĢ£ Ļ▓Į Ļ┤Ć ņĪ░Ļ▓Į ļ░Å ņ¢┤ļ®öļŗłĒŗ░ ņ”Øņ¦ä ņ×ÉņøÉņ£╝ļĪ£ņØś ĒÖ£ņÜ® Ļ░ĆļŖźņä▒ ļśÉĒĢ£ ĻĖ░ļīĆļÉ£ļŗż.
ņĀüņÜö
ļ│Ė ņŚ░ĻĄ¼ļŖö ļīĆĒæ£ņĀü ĒŚłļĖīņŗØļ¼╝ļĪ£ ņĢīļĀżņĀĖ ņ׳ļŖö ļ»╝ĒŖĖļźśņØś ņŚ╝ņŖżĒŖĖļĀłņŖż ņŚÉ ļīĆĒĢ£ ļ░śņØæ ĻĖ░ņ×æņØä ĻĄ¼ļ¬ģĒĢśĻĖ░ ņ£äĒĢ£ ļ¬®ņĀüņ£╝ļĪ£ NaClņØä ņ▓śļ”¼ĒĢśņŚ¼ ļ»╝ ĒŖĖņØś ņāØņ£Ī ļ░Å ņāØļ”¼ņĀü ļ░śņØæņØä ņĪ░ņé¼ĒĢśņśĆļŗż. ĒÄśĒŹ╝ļ»╝ĒŖĖ, ņśżļŹ░ņĮöļĪ▒ļ»╝ĒŖĖ, ņ┤łņĮöļ»╝ĒŖĖ, ņŖżĒö╝ņĢäļ»╝ĒŖĖ, ņĢĀĒöīļ»╝ĒŖĖ, ĒīīņØĖņĢĀĒöīļ»╝ĒŖĖ ļō▒ 6Ļ░Ćņ¦Ć ļ»╝ĒŖĖļźśņØś ņāØņ£Ī ļ░Å ņāØļ”¼ņĀü ļīĆņé¼ļ░śņØæņØä ņĪ░ņé¼ ļ╣äĻĄÉļČäņäØĒĢ£ Ļ▓░Ļ│╝ ņŚ╝ ņ▓śļ”¼ ļåŹļÅäĻ░Ć ņ”ØĻ░ĆļÉĀņłśļĪØ ļ»╝ĒŖĖļźśņØś ņ┤łņן, ņŚĮņן, ņŚĮĒÅŁņØĆ ņżäņ¢┤ļōĀ ļ░śļ®┤, ņ¦ĆņāüļČĆ ņāØņ▓┤ ņżæņØĆ ņĀĆļåŹļÅä ņŚ╝ ņ▓śļ”¼ņŚÉņä£ļŖö ļīĆņĪ░ĻĄ¼ļ│┤ļŗż ņĢĮĻ░ä ņ”ØĻ░ĆļÉśļŖö Ļ▓āņ£╝ļĪ£ ļéśĒāĆ ļé¼ļŗż. ļśÉĒĢ£ ļ»╝ĒŖĖ ĒÆłņóģ ļ│äļĪ£ ņ░©ņØ┤Ļ░Ć ņ׳ĻĖ┤ ĒĢśņśĆņ¦Ćļ¦ī ņØ╝ņĀĢ ņŚ╝ ļåŹļÅäĻ╣īņ¦Ć ļŖö ĒĢŁņé░ĒÖö ĒÖ£ņä▒, ņ┤Ø ĒÅ┤ļ”¼ĒÄśļåĆ, ņ┤Ø ĒöīļØ╝ļ│┤ļģĖņØ┤ļō£ ĒĢ©ļ¤ēņØ┤ ļīĆņĪ░ĻĄ¼ļ│┤ļŗż ņ”ØĻ░ĆļÉśļŖö Ļ▓āņØä ņĢī ņłś ņ׳ņŚłļŗż. ļé┤ņŚ╝ņä▒ņØ┤ Ļ░ĢĒĢśļ®┤ņä£ ĒĢŁņé░ĒÖö ĒÖ£ņä▒, ņ┤Ø ĒÅ┤ ļ”¼ĒÄśļåĆ, ņ┤Ø ĒöīļØ╝ļ│┤ļģĖņØ┤ļō£ ĒĢ©ļ¤ēņØ┤ ļ¦ÄņØĆ ĒÆłņóģņØĆ ņ┤łņĮöļ»╝ĒŖĖņÖĆ ņŖżĒö╝ņĢäļ»╝ ĒŖĖņØ┤ļ®░, ņŖżĒö╝ņĢäļ»╝ĒŖĖļŖö ņŚ╝ ļåŹļÅäņŚÉ ļö░ļØ╝ ņ░©ņØ┤Ļ░Ć ļÜ£ļĀĘĒĢśņśĆļŗż. ļ░śļ®┤ņŚÉ ņŚ╝ņŚÉ ļ»╝Ļ░ÉĒĢśļ®┤ņä£ ĒĢŁņé░ĒÖö ĒÖ£ņä▒, ņ┤Ø ĒÅ┤ļ”¼ĒÄśļåĆ, ņ┤Ø ĒöīļØ╝ļ│┤ļģĖņØ┤ļō£ ĒĢ©ļ¤ē ņØ┤ ļé«ņØĆ ĒÆłņóģņØĆ ņĢĀĒöīļ»╝ĒŖĖļĪ£ ļéśĒāĆļé¼ļŗż.




