

An Analysis of the Growth and Photosynthetic Responses of Potted Veronica pusanensis Y.N.Lee according to the Shading Levels
-
ParkSe Hoon
 , LeeJae Hwan, NamSang Yong
, LeeJae Hwan, NamSang Yong
- Received April 17, 2023; Revised May 16, 2023; Accepted May 23, 2023;
- ABSTRACT
-
- Background and objective: Veronica pusanensis, an endemic species of South Korea belonging to the Plantaginaceae family, is found in Busan, South Korea. Due to its high ornamental value, it is expected to be utilized as a flower crop. However, currently it is an endangered species with its habitats being destroyed and reduced.
- Methods: Therefore, in this study, we analyzed the growth and photosynthetic responses of potted plants of V. pusanensis under different shading levels to enable mass production. Polyethylene (PE) shading films were selected as the shading material, and shading levels were designed at 0, 35, 45, 60, 75, and 99%, respectively.
- Results: Results showed that shoot height, shoot width, ground cover, leaf length, leaf width, shoot fresh and dry weight, chlorophyll content (SPAD units), and chlorophyll fluorescence parameters Fv/Fm and PIABS were highest under the 35% shading level. This indicates that it is relatively more desirable to grow V. pusanensis in shade culture compared to under direct sunlight. Meanwhile, root fresh weight and dry weight were highest under the 0% shading level.
- Conclusion: Therefore, it is recommended to grow V. pusanensis under direct sunlight to significantly increase root biomass when the purpose is to facilitate rootage when transplanting plants for habitat restoration. On the other hand, for the cultivation of V. pusanensis as an ornamental flower crop, it is recommended to grow the plants under the 35% shading level to significantly increase plant sizes and maintain ensure the proper functioning of photosynthetic responses in photosystem II (PSII).
- Introduction
- Introduction
Veronica was previously known to belong to the Scrophulariaceae family, but now belongs to the Plantaginaceae family (Olmstead, 2002). Veronica is distributed throughout the Northern Hemisphere and the Australasian region, and various species are discovered also in Western Asia and New Zealand (Albach et al., 2004). Approximately 450 species of Veronica are distributed worldwide (Albach et al., 2008), and they grow naturally in broad and diverse regions from coastal to alpine areas due to their excellent ecological adaptability (Albach et al., 2005). Veronica plants live in diverse environments such as terrestrial, semi-aquatic, and aquatic depending on the species, showing excellent adaptability (Salehi et al., 2019). In terms of medicinal use, extracts of Veronica plants are known to be effective in eliminating free radicals (Harput et al., 2011), have anti-inflammatory effects, and show cytotoxic activity in cancer cells such as KB epidermoid carcinoma and B16 melanoma (Harput et al., 2002). Moreover, Veronica plants also showed an effect in treating rheumatism (Beara et al., 2015), lung and respiratory diseases (Grundemann et al., 2013), and antithrombosis and wounds (Küpeli et al., 2005), thereby expected to be used effectively as a medicinal crop.V. pusanensis is a rare plant found in Busan, South Korea and is an endangered species that needs protection because its habitats are decreasing and it is damaged by poaching (Shin et al., 2012). Since V. pusanensis grows as a creeping habit and tends to cover the ground, it is expected to be used as a ground cover plant in coastal areas or gardens (Song et al., 2020a), and it can also be supplied as flower crops if they can be mass-produced since it has excellent ornamental value. Various physiological studies have been conducted in the past, such as a cuttage study using auxin in V. pusanensis (Kim et al., 2021), an investigation on salinity tolerance (Kwon et al., 2022), a study on light intensity treatment (Song et al., 2020a), a physiological exploration on the growth and flowering response according to photoperiod and temperature treatment (Song et al., 2020b), and a study on sprouting response according to temperature treatment (Song et al., 2019). However, there is insufficient technology for mass production and protection of the plant of V. pusanensis from abiotic stress during the cultivation period. As previously mentioned, V. pusanensis is an endangered species with a small wild population and rapid habitat destruction. Thus, by establishing the cultivation technology for mass production, it would be possible to help restore the habitats and also supply this species as a flower crop.Potted cultivation has a relatively small soil volume and soil moisture content due to the limited pot capacity and thus has high requirements for management compared to open-field cultivation (Lee and Nam, 2022b). However, unlike open-field cultivation, potted cultivation allows plant movement and facilitates the control of plant growth. Commercial plastic pots are typically brown or black, and these colors can significantly increase soil temperature during intense summer sunlight, leading to moisture loss and causing complex abiotic stress for plants (Lee and Nam, 2022b). Therefore, it is necessary to come up with methods to protect plans such as shading.To induce the stable growth of plants, newly planted or cutting plants must not be exposed to direct sunlight, and the plant should be protected from rapid environmental changes by using shading materials such as shading nets or shading films (Fowler and Chaffee, 2010; Semchenko et al., 2012). Plants grown under adequate shading levels are protected from rapid temperature rise, photodamage, and lack of soil moisture, and can be managed in a relatively stable environment (Nam et al., 2022). Moreover, since they are relatively low-maintenance and have a long irrigation cycle, they also help reduce labor during cultivation (Nam et al., 2022). Depending on the plant species, the plant cells might be damaged when exposed to strong direct sunlight, which causes the chloroplasts on the cell surface to move to the side walls, and this is referred to as chloroplast avoidance movement (Kasahara et al., 2002). Chlorophyll fluorescence analysis is one of the methods to identify plant stress levels and the inactivity of the reaction center of photosystem II (PSII), which can be done in the non-destructive investigation (Lechaudel et al., 2010; Lee et al., 2022b; Shim and Jeon, 2022). Chlorophyll fluorescence analysis that had been applied in various studies in the past had its usefulness proved by being used in assessing the photosynthetic efficiency and stress of plants through various fluorescence parameters such as Fv/Fm, ΦDo, ABS/RC, DIo/RC, and PIABS (An et al., 2022; Choi et al., 2022; Kwon et al., 2021; Lee et al., 2022b; Oh and Lee, 2022; Yang et al., 2022).Accordingly, this study established the fundamental data on the growth and photosynthetic responses of V. pusanensis affected by shading levels to contribute to the mass production of V. pusanensis.
- Research Methods
- Research Methods
- Plant material
- Plant material
As the plant material, we selected Veronica pusanensis which is an endemic species of South Korea and has high ornamental value. We selected plants around the shoot height of 3 cm and shoot width of 5 cm and applied them to the study.- Experimental environment and shading levels
- Experimental environment and shading levels
The study was conducted for a total of 5 weeks from May 9 to June 14, 2022, at the experimental greenhouse (37°38′40″ N 127°06′25″ E) in the Department of Environmental Horticulture, Sahmyook University. Plants were planted in round plastic pots with a diameter and height of 11 × 10.5 cm, at the center of the pot. Here, we used horticultural substrates containing fertilizers (Hanareumsangto, Shinsung Mineral, South Korea). We designed six shading levels: no shade (i.e., direct sunlight) (0%), greenhouse glass (35%), greenhouse glass and 1 layer of clear polyethylene (PE) shading film (45%), greenhouse glass and 1 layer of white PE shading film (60%), greenhouse glass and 2 layers of white PE shading film (75%), and greenhouse glass and 1 layer of black PE shading film (99%). For the photosynthetic photon flux density (PPFD) of each shading level, we calculated the mean and standard deviation of the measurements taken at 1 p.m. once a week on sunny days using a portable spectral radiometer (SpectraPen mini, Photon Systems Instruments, Czech Republic) (Table 1).The plants of all treatments including the control (0% shading level) were arranged on beds that are 1 m high, and the average temperature in the greenhouse during the study was 20.9 ± 2.1°C, and the average temperature of the outdoor area where the control was placed was 20.1 ± 2.0°C (Fig. 1A). Here, the relative humidity in the green-house was 58.5 ± 9.5%, and the relative humidity of the outdoor area was 60.0 ± 9.6% (Fig. 1B). The average cloud cover was 4.6 ± 2.2 okta, and this was evaluated by visual observation, which is the same as the measurement method of Korea Meteorological Administration, at 1 p.m. every day (Fig. 1C). The plants were watered 3 times a week. In this study, watering continued until gravitational water drainage occurred.- Measurement methods for growth and leaf color parameters
- Measurement methods for growth and leaf color parameters
We measured shoot height, shoot width, root length, shoot fresh and dry weight, root fresh and dry weight, ground cover, leaf length, leaf width, and leaf color of CIELAB L*, a*, and b* values. Here, we measured shoot height from the soil surface in the pot up to the point where the plant is the tallest and shoot width by measuring the broadest area of the plant seen from above. For root length, we measured the longest of the plant roots. Secondary analysis was conducted for ground cover by squaring the shoot width of the plant width. Fresh weight was measured after washing the plant and naturally drying it for 12 hours in an enclosed space, and dry weight was measured after hot air drying of the plant for 24 hours using a hot air dryer (HK-DO135F, HANKUK S&I, South Korea). CIELAB values (L*, a*, and b*) were measured by setting the spectrophotometer (CM-2600d, Konica Minolta, Japan) to CIELAB D65/10° with reference to the leaf color measurement method Lee et al. (2022a), after which we collected data including specular components. For RHS values, we compared the results of each of L*, a*, and b* with the Royal Horticultural Society (RHS) colour charts edition V and evaluated the leaf colors by selecting 2 approximate RHS values for each treatment. In addition, for leaf colors, we converted the means of CIELAB L*, a*, and b* to converted colors using Converting Colors designed by Zettl (2023).- Measurement methods for chlorophyll fluorescence
- Measurement methods for chlorophyll fluorescence
We analyzed the chlorophyll content and photosynthetic state of V. pusanensis affected by shading levels. Here, we measured the chlorophyll content using a portable chlorophyll meter (SPAD-502, Konica Minolta, Japan) and estimated the chlorophyll content relative to the same area for each treatment using SPAD units. We measured chlorophyll fluorescence responses using a portable chlorophyll fluorometer (FluorPen FP 110/D, Photon Systems Instruments, Czech Republic) by measuring the part where the leaf veins do not pass among the central part of the fully developed leaf. Here, before measuring the chlorophyll fluorescence, the plant was dark-adapted for approximately 15 minutes using dark-adaptation leaf clips according to the manufacturer’s guidelines (PSI, 2023), and we measured 5 times each in 3 replications on June 14, 2022, which is the end date of the study. We selected and applied five chlorophyll fluorescence parameters that can be used as plant stress indicators. We used the following equations to calculate Fv/Fm (1) representing the maximum quantum yield of photosystem II (PSII), ΦDo (2) representing the probability that absorbed photons will be dissipated, ABS/RC (3) representing absorption flux per reaction center, DIo/RC (4) representing dissipated energy flux per reaction center, and PIABS (5) that represents the performance index of absorption basis (PSI, 2023; Stirbet and Govindjee, 2011).- Statistical analysis
- Statistical analysis
Analysis of variance (ANOVA) was conducted using SAS 9.4 (SAS Institute, USA) to analyze the results of the study. To compare the means, statistical analysis was conducted using Duncan’s multiple range test at p < .05. The study was in a completely randomized design, with 5 plants assigned to each treatment in 3 replications.
- Results and Discussion
- Results and Discussion
- Analysis of plant sizes, biomass, and leaf color
- Analysis of plant sizes, biomass, and leaf color
Shading levels had diverse effects on the plant sizes of Veronica pusanensis (Fig. 2). The results showed that shoot height of V. pusanensis was highest at 14.78 cm under the 35% shading level (Fig. 3A). Sedum zokuriense showed the highest shoot height under the 65% shading level (Lee et al., 2021b), whereas Sarcandra glabra showed the highest shoot height under the 50% shading level and Ardisia crenata under the 35% shading level (Bae et al., 2016), indicating that the preferred shading levels vary among species. Meanwhile, shoot width was widest at 29.21 and 28.84 cm under the 35 and 45% shading levels, indicating that shade culture is better than cultivation under direct sunlight in order to significantly increase the plant shoot sizes (Fig. 3B). Root length was longest at 21.87 cm under the 60% shading level (Fig. 3C). A previous study reported that root length of V. nakaiana and V. pyrethrina was in inverse proportion to the increase of shading levels (Kwon et al., 2020). On the contrary, in this study, root length increased along with shading levels from 0% to 60%, showing conflicting results from a previous study, but the results from 75% to 99% shading levels were similar to a previous study. Like the results of shoot width, ground cover was high at 868.1 and 861.8 cm2 under the 35 and 45% shading levels (Fig. 3D). Leaf length was highest at 3.57 cm under the 35% shading level (Fig. 3E). Meanwhile, leaf width was highest at 3.46 cm under the 35% shading level, which was similar to the results of shoot height, shoot width, and leaf length (Fig. 3F). A previous study showed similar results as this study with Litsea japonica showing the highest leaf length and leaf width under the 35% shading level, but Osmanthus fragrans var. aurantiacus and Pittosporum tobira showed the highest leaf length and leaf width under the 50% shading level (Choi et al., 2012a), indicating that the responses to leaf growth according to shading levels vary among species. Phenotypic plasticity is the ability of a plant to change its morphological characteristics so that it can adapt to the changing growth environment (Bradshaw, 1965; DeWitt et al., 1998; Sultan, 1987). Among phenotypic plasticity, shade-induced phenotype induces the plant to show the morphological characteristics suitable to shade (Weijschede et al., 2006). In a shade-induced phenotype, we can expect to see an increase in the plant’s shoot height, shoot width or ground cover, and leaf sizes (Lee and Nam, 2022b), and this change is expected to help buffer the negative impact on growth due to the reduced amount of light received in shade.Shoot fresh weight was highest at 18.51 g under the 35% shading level (Fig. 4A). Meanwhile, V. rotunda var. subintegra showed the highest shoot fresh weight when grown under the 55% shading level in the seedling process (Lee et al., 2020b). Root fresh weight was highest at 3.53 g under the 0% shading level (Fig. 4B). Shoot dry weight was highest at 2.58 g under the 35% shading level (Fig. 4C). Similarly, stem dry weight of Illicium anisatum was highest under the 35% shading level (Choi et al., 2012b). Root dry weight was highest at 0.87 g under the 0% shading level (Fig. 4D). Both root fresh weight and dry weight were highest under the 0% shading level, which was contrary to the result that root length was highest under the 60% shading level. This may be because factors such as carbohydrate content and thickness in the main root increased while the length of the side roots relatively decreased as there was a greater amount of light received. In addition, stimulation by water stress may have also caused an increase in the root fresh weight and dry weight. In previous studies, root fresh weight and dry weight of V. nakaiana and V. pyrethrina (Kwon et al., 2020) and V. rotunda var. subintegra were highest under the 0% shading level (Lee et al., 2020a), and root dry weight of Distylium racemosum, Neolitsea aciculata, and Stauntonia hexaphylla was also highest under the 0% shading level, showing consistent results with this study (Choi et al., 2012b). In summary of the results of this study and previous studies, shading was better than direct sunlight to significantly increase shoot biomass. On the other hand, to significantly increase root biomass, it was relatively more beneficial to grow under direct sunlight. In addition, in the plant sizes and biomass analysis, the control (0% shading level) was placed outside the greenhouse and shading film due to limitations in the study, and thus it could have been affected by additional abiotic stress due to air temperature and soil temperature rise, humidity change, and damage caused by rainfall. In particular, as shown in the results, there may have been a negative effect on shoot growth, which is something to refer to.CIELAB is the color space made in 1976 by the International Commission on Illumination (Commission Internationale de l’Eclairage; CIE) in France (Lee et al., 2022c). CIELAB was developed in 1976 to make up for the deficiencies of Hunter Lab developed in 1958, and unlike Hunter Lab indicated in L, a, and b, it is indicated in L*, a*, and b* that include asterisk (Lee and Nam, 2022a). In this study, the leaf color analysis of V. pusanensis using CIELAB showed different results by shading level (Table 2). CIELAB L*, which is the color space coordinate representing lightness, was highest at 46.90 under the 0% shading level. Meanwhile, CIELAB a* that represents green-red opponent colors showed the same significance level under the 35–60% shading levels. CIELAB b* that represents blue-yellow opponent colors was highest at 24.64 under the 0% shading level. b* tended to be relatively high in the treatment where growth parameters such as shoot height, shoot width, fresh weight, and dry weight were low. The results of previous studies showed some similarities with this study, proving that the growth rates of Orostachys japonica and O. boehmeri (Lee et al., 2022c), and Delosperma cooperi (Lee et al., 2022d) had a negative correlation with b*. Meanwhile, L* and b* showed a positive correlation with each other (Kim et al., 2022; Lee et al., 2022c; 2022d). Similarly, this study also showed that L* and b* increased together under the 0% shading level, implying that it would be possible to use L* and b* as stress indicators for the leaf color of some plants including V. pusanensis. Royal Horticultural Society (RHS) values were 146B and 147B under the 0% shading level, showing that the color was relatively closer to yellow. On the other hand, the values were 137A and N137D under the 35–60% shading levels, showing that the leaf color was relatively green.As a result, assuming that increased plant sizes and dark green leaf color contribute positively to the purchasing power of consumers when supplying V. pusanensis as an ornamental flower crop, it is recommended to grow V. pusanensis under the 35% shading level in order to significantly increase plant sizes and maintain dark green leaf color.- Analysis of chlorophyll content and fluorescence
- Analysis of chlorophyll content and fluorescence
Chlorophyll content (SPAD units) was highest in the order of 35 to 45% shading levels at 50.94 and 50.55, respectively, showing the same results as shoot width and ground cover (Fig. 5A). This is consistent with the results of previous studies that, when growing Hylotelephium telephium (Nam et al., 2022), Hoya carnosa and Spathiphyllum wallisii (Lee et al., 2021a), and S. zokuriense (Lee et al., 2021b) grown under conditions of optimal shading levels and light intensity, an increase in chlorophyll content was observed concomitant with high shoot width. Meanwhile, V. rotunda var. subintegra showed relatively higher chlorophyll content under the 55–75% shading levels compared to the 0% shading level (Kwon et al., 2020). In summary of the results above, some plants including V. pusanensis seem to have homeostasis to maintain a constant total amount of carbon dioxide assimilation by maximizing plant sizes and chlorophyll density under low light intensity. Fv/Fm, which represents maximum quantum yield of photosystem II (PSII), was highest at 0.85 under the 35% shading level, and lowest at 0.81 under the 99% shading level (Fig. 5B). Fv/Fm, which is a parameter that represents a maximum quantum yield of PSII, is known to range from 0.78 to 0.84 in higher plants species that are not stressed (Björkman and Demmig, 1987; Butler and Kitajima, 1975; Genty et al., 1989; Govindjee, 1995; Govindjee, 2004; Paillotin, 1976; Yoo et al., 2012). Fv/Fm was slightly higher than the normal range under the 35% shading level, whereas it was within the normal range in other treatments. In particular, the maximum quantum yield was significantly low under the 99% shading level where the light intensity was extremely low, but it did not have a negative effect so much as to significantly deteriorate the function of the reaction center of PSII. As a result, plant sizes or biomass of V. pusanensis was relatively low under the 99% shading level, but shade tolerance was excellent, and growth rates are expected to be quickly recovered by moving the plant to an environment with high light intensity. ΦDo that represents the probability that the absorbed photon will be dissipated was highest at 0.18 under the 99% shading level (Fig. 5C). ABS/RC that represents absorption flux per reaction center was highest at 1.99 under the 99% shading level (Fig. 5D). ABS/RC, one of chlorophyll fluorescence parameters, represents the state of the reaction center that is inactivated as the number of reaction centers in reduction state increases in PSII (Spoustová et al., 2013). Meanwhile, DIo/RC that represents dissipated energy flux per reaction center was highest at 0.38 under the 99% shading level, indicating that the energy use efficiency has decreased (Fig. 5E). PIABS is a performance index with an absorption basis (Srivastava et al., 1999). This index represents the degree of photosynthetic activity and is a set of light energy absorbing capacity, electron transfer efficiency, and electron fixation efficiency of PSII (Thach et al., 2007). PIABS represents the overall vitality of the photosynthetic apparatus (Strasser et al., 2000; Živčák et al., 2008), and it is used as an index that represents the soundness of plants (Oukarroum et al., 2007). PIABS was highest at 8.13 under the 35% shading level (Fig. 5F). This result was consistent with the results of chlorophyll content, shoot fresh weight, and dry weight, indicating that higher PIABS leads to higher chlorophyll density and more carbon dioxide assimilation.In conclusion, as a result of comprehensively evaluating the plant sizes, biomass, leaf color, and photosynthetic responses analysis of V. pusanensis affected by shading levels, we discovered that shading levels significantly increase the size of V. pusanensis and maintain dark green leaf color. It is recommended to cultivate the plant under the 35% shading level to maintain the activity of PSII in the optimal state.
- Conclusion
- Conclusion
Veronica pusanensis is an endemic species belonging to the Plantaginaceae family and found in Busan, South Korea. V. pusanensis has high ornamental value and can be used as a flower crop, but it is currently an endangered species with its habitats being destroyed and reduced. Accordingly, this study analyzed the growth and photosynthetic responses of potted V. pusanensis according to shading levels for mass production of V. pusanensis. We selected polyethylene (PE) shading films as shading materials and designed 6 shading levels such as 0, 35, 45, 60, 75, and 99%, respectively. The results showed that shoot height, shoot width, ground cover, leaf length, leaf width, shoot fresh weight and dry weight, chlorophyll content (SPAD units), and chlorophyll fluorescence parameters Fv/Fm, PIABS were highest under the 35% shading level, indicating that it is relatively more desirable to grow V. pusanensis in shade culture compared to under direct sunlight. Meanwhile, root fresh weight and dry weight was highest under the 0% shading level, indicating that it is recommended to grow under direct sunlight to significantly increase root biomass when the purpose is to facilitate rootage when transplanting plants for habitat restoration. To cultivate V. pusanensis as an ornamental flower crop, it is recommended to grow under the 35% shading level to significantly increase plant sizes and maintain ensure the proper functioning of photosynthetic responses of photosystem II (PSII).
Fig. 1
Temperature, relative humidity, and cloud cover in this study. Cloud cover index, 0 okta: sky clear; 1–2 okta: few clouds; 3–4 okta: scattered; 5–7 okta: broken; 8 okta: overcast; 9 okta: sky obscured.
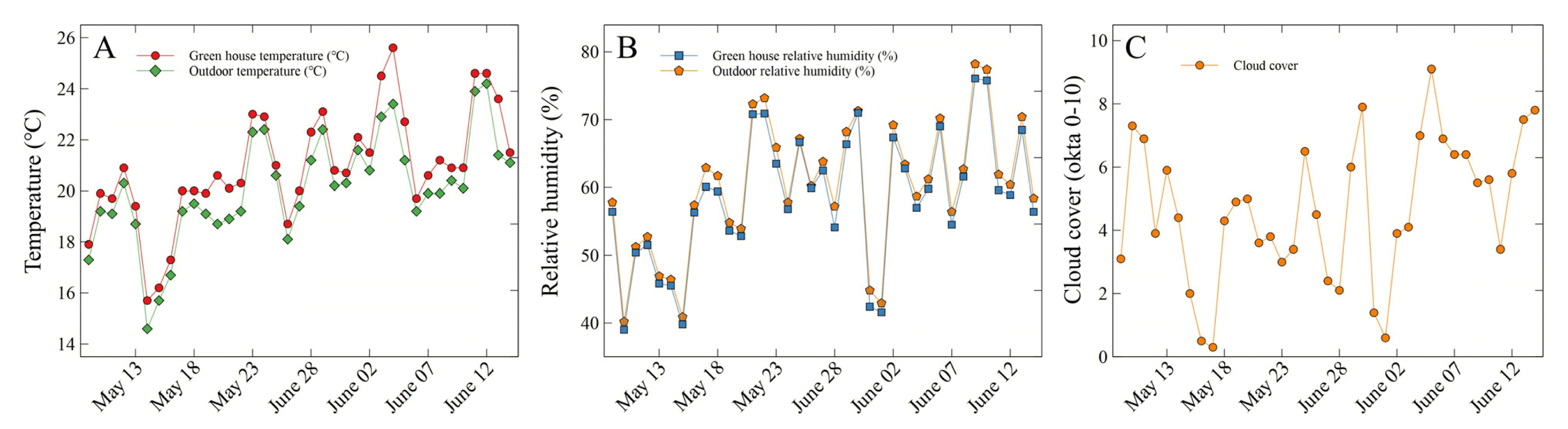

Fig. 3
Plant sizes of V. pusanensis as affected by shading levels for 5 weeks. Vertical bars indicate the standard error, and asterisks (***) indicate significant at p < .001. Different lowercase letters indicate significant differences at p < .05 based on Duncan’s multiple range test (DMRT).
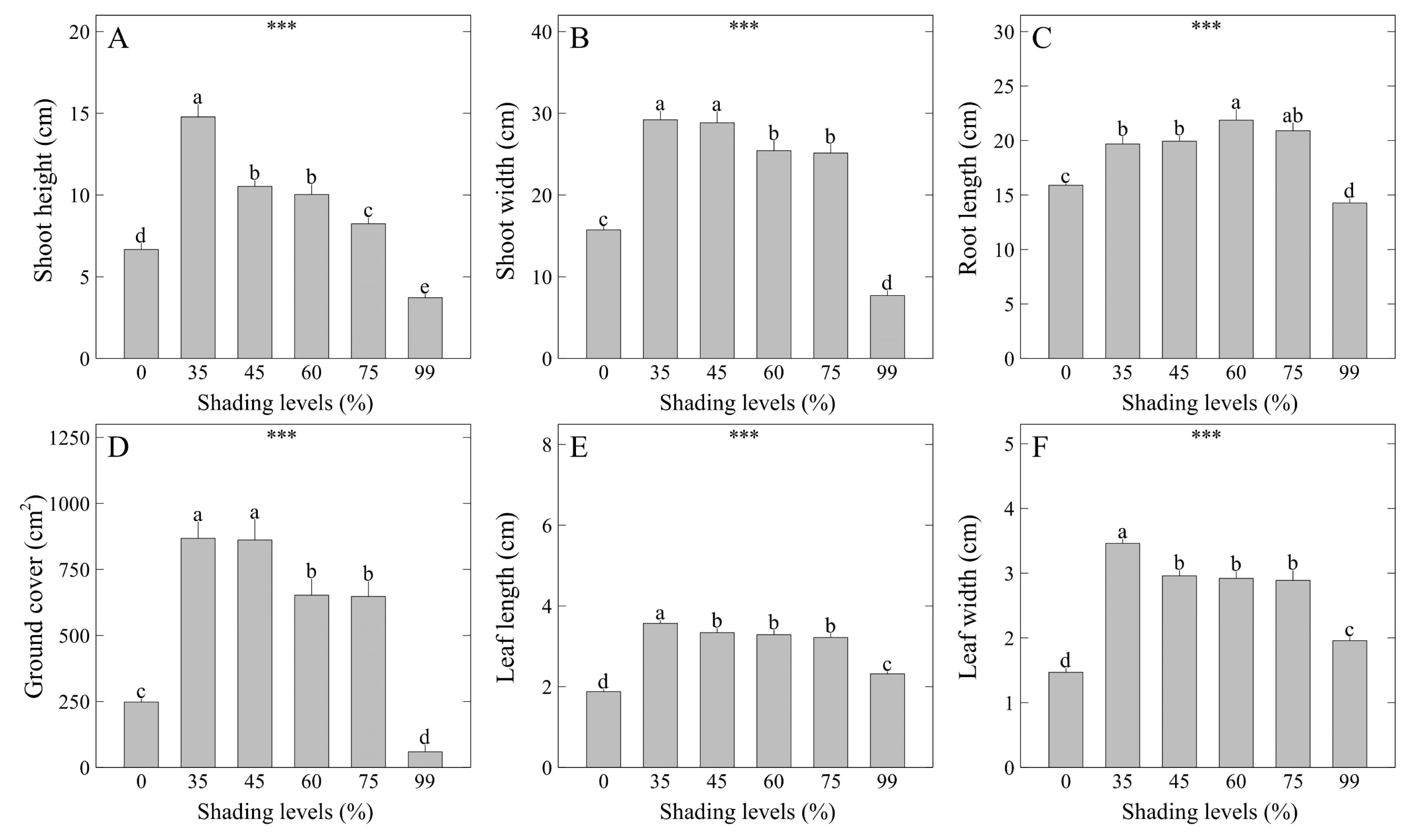
Fig. 4
Shoot and root weight of V. pusanensis as affected by shading levels for 5 weeks. Vertical bars indicate the standard error, and asterisks (***) indicate significant at p < .001. Different lowercase letters indicate significant differences at p < .05 based on DMRT.
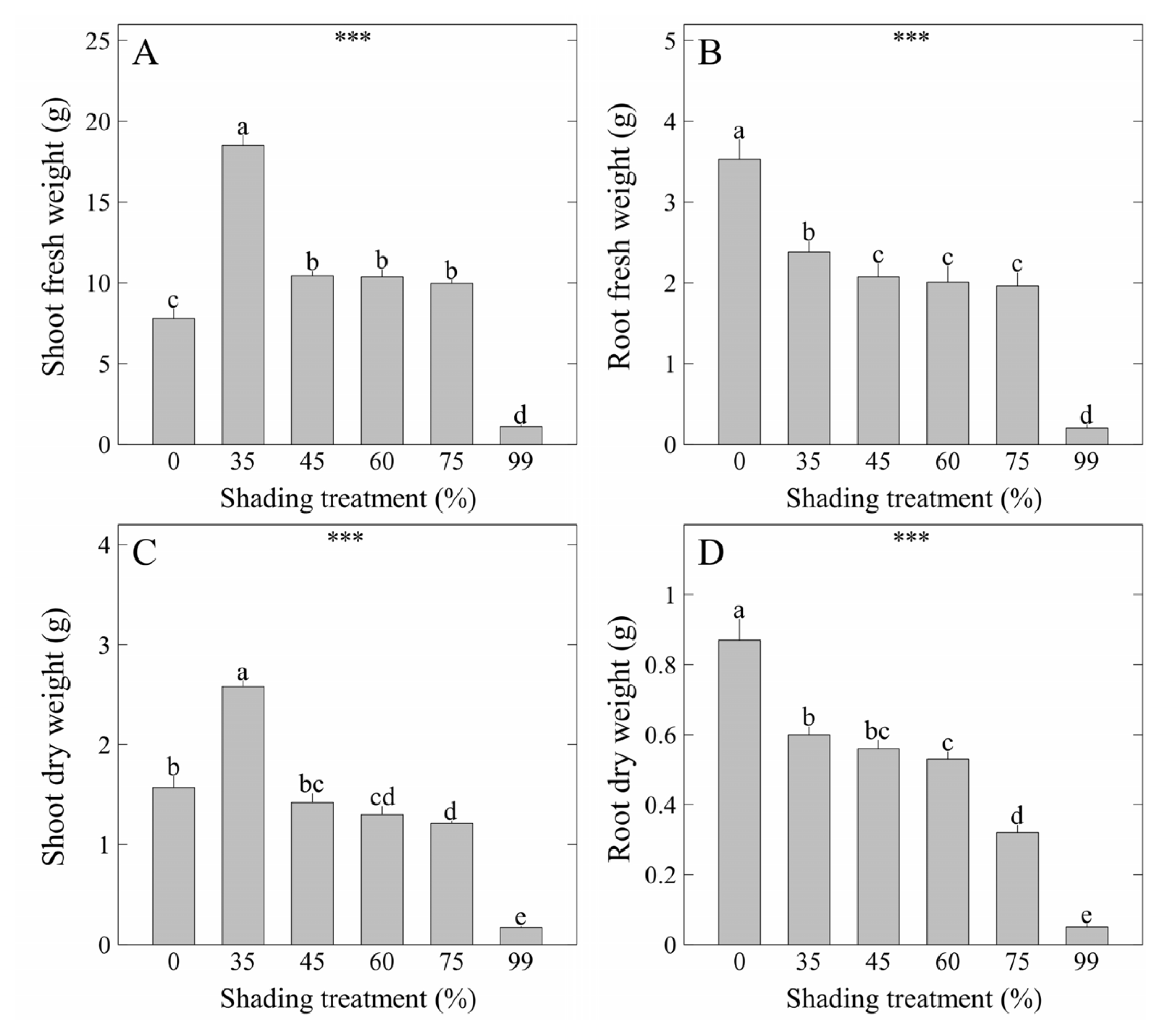
Fig. 5
Cholophyll content (SPAD units) and photosynthetic responses of V. pusanensis as affected by shading levels for 5 weeks. Vertical bars indicate the standard error, and asterisks (***) indicate significant at p < .001. Different lowercase letters indicate significant differences at p < .05 based on DMRT.
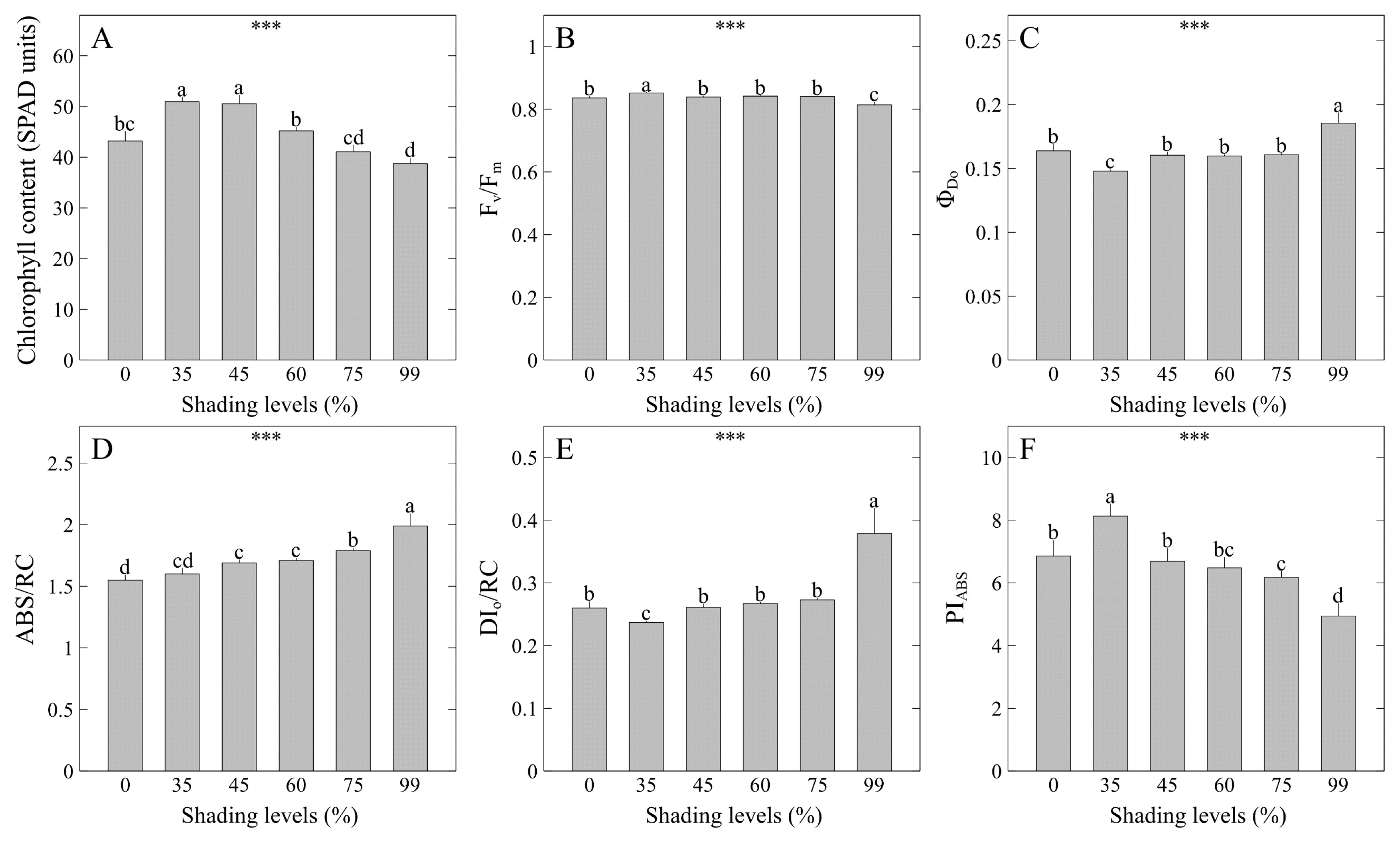
Table 1
Shading levels of photosynthetic photon flux density (PPFD) in this study
| Standard shading levels (%) | Relative shading levels (%) | PPFD (μmol m−2 s−1) |
|---|---|---|
| 0 | 0 | 1783.2 ± 193.2z |
| 35 | 36.1 | 1140.9 ± 143.4 |
| 45 | 44.2 | 996.1 ± 121.6 |
| 60 | 60.8 | 699.8 ± 94.8 |
| 75 | 74.1 | 460.6 ± 58.0 |
| 99 | 99.1 | 16.3 ± 4.9 |
Table 2
Leaf color reading values of CIELAB values (L*, a*, b*), RHS values, and converted color of Veronica pusanensis as affected by shading levels for 5 weeks
| Shading levels (%) | CIELAB values | RHS valuesz | Converted colory (color chip) | ||
|---|---|---|---|---|---|
|
|
|||||
| L* | a* | b* | |||
| 0 | 46.90 ax | −10.49 c | 24.64 a | 146B, 147B |

|
| 35 | 38.91 c | −8.51 a | 16.98 c | 137A, N137D |

|
| 45 | 39.73 c | −8.39 a | 17.21 c | 137A, N137D |

|
| 60 | 38.89 c | −8.35 a | 17.43 c | 137A, N137D |

|
| 75 | 41.74 b | −9.11 b | 21.89 b | 146A, 148A |

|
| 99 | 42.85 b | −9.52 b | 21.02 b | 146A, 148A |

|
|
|
|||||
| Significancew | *** | *** | *** | ||
- REFERENCES
- REFERENCES
References
Albach, D.C., H.M. Meudt, B. Oxelman. 2005. Piecing together the “new” Plantaginaceae. America Journal of Botany. 92(2):297-315. https://doi.org/10.3732/ajb.92.2.297
[Article]Albach, D.C., M.M. Martinez-Ortega, L. Delgado, H. Weiss-Schneeweiss, F. Özgökce, M.A. Fischer. 2008. Chromosome numbers in Veroniceae (Plantaginaceae): review and several new counts. Annals of the Missouri Botanical Garden. 95(4):543-566. https://doi.org/10.3417/2006094
[Article]Albach, D.C., M.M. Martinez-Ortega, M.A. Fischer, M.W. Chase. 2004. A new classification of the tribe Veroniceae—problems and a possible solution. Taxon. 53(2):429-452. https://doi.org/10.2307/4135620
[Article]An, H.R., S.Y. Lee, P.M. Park, Y.J. Kim. 2022. Effects of substrate water contents on post-shipping performance of Phalaenopsis . Horticultural Science and Technology. 40(1):52-62. https://doi.org/10.7235/HORT.20220006
[Article]Bae, E.J., E.J. Jin, J.H. Bae, K.S. Lee, S.M. Choi. 2016. Growth and physiological characteristics of Sarcandra glabra and Ardisia crenata under different light intensity. Journal of Korean Society for People, Plants, and Environment. 19(2):85-93. https://doi.org/10.11628/ksppe.2016.19.2.85
[Article]Beara, I., J. Živković, M. Lesjak, J. Ristić, K. Šavikin, Z. Maksimović, T. Janković. 2015. Phenolic profile and anti-inflammatory activity of three Veronica species. Industrial Crops and Products. 63:276-280. https://doi.org/10.1016/j.indcrop.2014.09.034
[Article]Björkman, O., B. Demmig. 1987. Photon yield of O2 evolution and chlorophyll fluorescence characteristics at 77 K among vascular plants of diverse origins. Planta. 170:489-504. https://doi.org/10.1007/BF00402983
[Article] [PubMed]Bradshaw, A.D. 1965. Evolutionary significance of phenotypic plasticity in plants. Advances in Genetics. 13:115-155. https://doi.org/10.1016/S0065-2660(08)60048-6
[Article]Butler, W.L., M. Kitajima. 1975. Fluorescence quenching in photosystem II of chloroplasts. Biochimica et Biophysica Acta. 376(1):116-125. https://doi.org/10.1016/0005-2728(75)90210-8
[Article] [PubMed]Choi, D.S., T.K.L. Nguyen, M.M. Oh. 2022. Growth and biochemical responses of kale to supplementary irradiation with different peak wavelengths of UV-A light-emitting diodes. Horticulture, Environment, and Biotechnology. 63(1):65-76. https://doi.org/10.1007/s13580-021-00377-4
[Article]Choi, S.M., H.C. Shin, K.S. Lee, E.G. Bae, K.O. Choi, K.Y. Huh. 2012a. Effects of shading rates on growth characteristics and photosynthesis in four broad-leaved evergreen trees. Journal of Korean Society for People, Plants, and Environment. 15(2):99-106.Choi, S.M., H.C. Shin, K.Y. Huh, H.J. Jung. 2012b. Seedling quality of broad-leaved evergreen trees with different shading levels. Journal of Korean Society for People, Plants, and Environment. 15(4):265-271.DeWitt, T.J., A. Sih, D.S. Wilson. 1998. Costs and limits of phenotypic plasticity. Trends in Ecology and Evolution. 13(2):77-81. https://doi.org/10.1016/S0169-5347(97)01274-3
[Article] [PubMed]Fowler, J., L. Chaffee. 2010. General information on propagation by stem cuttings University of California Davis. California, USA:Genty, B., J.M. Briantais, N.R. Baker. 1989. The relationship between the quantum yield of photosynthetic electron transport and quenching of chlorophyll fluorescence. Biochimica et Biophysica Acta (BBA) - General Subjects. 990(1):87-92. https://doi.org/10.1016/S0304-4165(89)80016-9
[Article]Govindjee, G. 1995. Sixty-three years since Kautsky: chlorophyll a fluorescence. Australian Journal of Plant Physiology. 22(2):131-160.
[Article]Govindjee, G. 2004. Chlorophyll a fluorescence: a bit of basics and history. In: Papageorgiou G.C., Govindjee , (Eds), Chlorophyll a fluorescence: a signature of photosynthesis. Advances in Photosynthesis and Respiration (pp. 19:1-41). Dordrecht, The Netherlands: Springer.
[Article]Grundemann, C., M. Garcia-Kaufer, B. Sauer, E. Stangenberg, M. Könczöl, I. Merfort, M. Zehl, R. Huber. 2013. Traditionally used Veronica officinalis inhibits proinflammatory mediators via the NF-κB signalling pathway in a human lung cell line. Journal of Ethnopharmacology. 145(1):118-126. https://doi.org/10.1016/j.jep.2012.10.039
[Article] [PubMed]Harput, U.S., I. Saracoglu, M. Inoue, Y. Ogihara. 2002. Anti-inflammatory and cytotoxic activities of five Veronica species. Biological and Pharmaceutical Bulletin. 25(4):483-486. https://doi.org/10.1248/bpb.25.483
[Article] [PubMed]Harput, U.S., Y. Genc, N. Khan, I. Saracoglu. 2011. Radical scavenging effects of different Veronica species. Records of Natural Products. 5(2):100-107.Kasahara, M., T. Kagawa, K. Oikawa, N. Suetsugu, M. Miyao, M. Wada. 2002. Chloroplast avoidance movement reduces photodamage in plants. Nature. 420(6917):829-832. https://doi.org/10.1038/nature01213
[Article] [PubMed]Kim, H.J., J.H. Lee, J.H. Lee, M.S. Ko, S.Y. Nam. 2022. A study on indoor cultivation of Petrosedum rupestre and P.rupestre cv. Angelina using commercial white T5 LEDs. Journal of Agricultural, Life and Environmental Sciences. 34(3):354-367. https://doi.org/10.22698/jales.20220035
[Article]Kim, S.H., J.H. Kim, H.J. Oh, S.Y. Kim, G.U. Suh. 2021. Vegetative propagation of Veronica dahurica and Veronica pusanensis by stem cuttings with auxins. Rhizosphere. 17:100315. https://doi.org/10.1016/j.rhisph.2021.100315
[Article]Küpeli, E., U.S. Harput, M. Varel, E. Yesilada, I. Saracoglu. 2005. Bioassay-guided isolation of iridoid glucosides with antinociceptive and anti-inflammatory activities from Veronica anagallis-aquatica L. Journal of Ethnopharmacology. 102(2):170-176. https://doi.org/10.1016/j.jep.2005.05.042
[Article] [PubMed]Kwon, H.H., H.J. Oh, J.H. Kim, S.Y. Kim. 2021. Development of raising seedling technology for Veronica pyrethrina Nakai using plug trays. Journal of People, Plants, and Environment. 24(5):499-507. https://doi.org/10.11628/ksppe.2021.24.5.499
[Article]Kwon, H.H., H.J. Oh, W. Cho, Y.H. Kwon, S.H. Yang, S.Y. Kim. 2022. Growth and physiological responses of Pseudolysimachion pusanensis (Y. N. Lee) Y. N. Lee to NaCl treatment. Journal of People, Plants, and Environment. 25(2):133-141. https://doi.org/10.11628/ksppe.2022.25.2.133
[Article]Kwon, Y.H., H.H. Kwon, M. Gil, M.J. Jeong, S.Y. Kim, Y.H. Rhie. 2020. Growth of Veronica nakaiana and Veronica pyrethrina under different shading levels. Flower Research Journal. 28(4):331-339. https://doi.org/10.11623/frj.2020.28.4.12
[Article]Lechaudel, M., L. Urban, J. Joas. 2010. Chlorophyll fluorescence, a nondestructive method to assess maturity of mango fruits (cv. ‘Cogshall’) without growth conditions bias. Journal of Agricultural and Food Chemistry. 58(13):7532-7538. https://doi.org/10.1021/jf101216t
[Article] [PubMed]Lee, J.H., H.B. Kim, S.Y. Nam. 2022a. Evaluation of the growth and leaf color of indoor foliage plants under high temperature and continuous lighting conditions at different light intensity. Journal of Agricultural, Life and Environmental Sciences. 34(1):26-36. https://doi.org/10.22698/jales.20220004
[Article]Lee, J.H., H.J. Yoo, S.Y. Nam. 2022b. Chlorophyll fluorescence response of indoor foliage plants as affected by light intensity levels under high temperature and continuous lighting conditions. Journal of Agriculture and Life Science. 56(1):19-26. https://doi.org/10.14397/jals.2022.56.1.19
[Article]Lee, J.H., R.A.M. Cabahug, N.H. You, S.Y. Nam. 2021a. Chlorophyll fluorescence and growth evaluation of ornamental foliage plants in response to light intensity levels under continuous lighting conditions. Flower Research Journal. 29(3):153-164. https://doi.org/10.11623/frj.2021.29.3.05
[Article]Lee, J.H., S.Y. Nam. 2022a. Analysis of growth and leaf color changes of Sedum album cv. Athoum according to the spectral power distribution of several white LEDs. Flower Research Journal. 30(4):184-193. https://doi.org/10.11623/frj.2022.30.4.03
[Article]Lee, J.H., S.Y. Nam. 2022b. Effects of shading treatment on the growth and leaf color quality of potted Phedimus takesimensis cv. Atlantis. Journal of Agricultural, Life and Environmental Sciences. 34(3):413-424. https://doi.org/10.22698/jales.20220040
[Article]Lee, J.H., S.Y. Soh, H.J. Kim, S.Y. Nam. 2022c. Effects of LED light quality on the growth and leaf color of Orostachys japonica and O. boehmeri . Journal of Bio-Environment Control. 31(2):104-113. https://doi.org/10.12791/KSBEC.2022.31.2.104
[Article]Lee, J.H., S.Y. Soh, S.Y. Nam. 2022d. Growth evaluation of potted Delosperma cooperi (Hook. f.) L. Bolus to shading levels, potting media, and fertilization rates. Flower Research Journal. 30(1):1-9. https://doi.org/10.11623/frj.2022.30.1.01
[Article]Lee, J.H., Y.S. Lim, S.Y. Nam. 2021b. Optimization of shading levels, potting media, and fertilization rates on the vegetative growth of Sedum zokuriense Nakai. Flower Research Journal. 29(4):239-246. https://doi.org/10.11623/frj.2021.29.4.04
[Article]Lee, S.I., S.H. Yeon, J.S. Cho, C.H. Lee. 2020a. Growth assessment of the potted cultivation of Veronica rotunda var. subintegra (Nakai) T. Yamaz. Flower Research Journal. 28(1):14-20. https://doi.org/10.11623/frj.2020.28.1.03
[Article]Lee, S.I., S.H. Yeon, J.S. Cho, M.J. Jeong, C.H. Lee. 2020b. Optimization of cultivation conditions on effective seedlings of Veronica rotunda var. subintegra (Nakai) T. Yamaz. Protected Horticulture and Plant Factory. 29(2):181-188. https://doi.org/10.12791/KSBEC.2020.29.2.181
[Article]Nam, J.W., J.H. Lee, J.G. Lee, S.Y. Hwang, S.Y. Nam. 2022. Characteristics of growth and leaf color of Hylotelephium telephium cv. Lajos and H. sieboldii cv. Mediovariegatum as affected by shading levels. Flower Research Journal. 30(4):172-183. https://doi.org/10.11623/frj.2022.30.4.02
[Article]Oh, S.I., A.K. Lee. 2022. Effect of rooting promoter treatments on cutting and growth of Sedum takesimense . Horticultural Science and Technology. 40(1):12-20. https://doi.org/10.7235/HORT.20220002
[Article]Olmstead, R.G. 2002. Whatever happened to Scrophulariaceae. Fremontia. 30:13-22.Oukarroum, A., S.E. Madidi, G. Schansker, R.J. Strasser. 2007. Probing the responses of barley cultivars (Hordeum vulgare L.) by chlorophyll a fluorescence OLKJIP under drought stress and re-watering. Environmental and Experimental Botany. 60(3):438-446. https://doi.org/10.1016/j.envexpbot.2007.01.002
[Article]Paillotin, G. 1976. Movement of excitations in the photosynthetic domains of photosystem II. Journal of Theoretical Biology. 58(1):237-252. https://doi.org/10.1016/0022-5193(76)90150-8
[Article]Photon Systems Instruments (PSI). 2023 April 17 FluorPen FP 110 PAR-FluorPen FP 110 Monitoring Pen MP 100. FluorPen & PAR FluorPen, Photon Systems Instruments website Retrieved from https://handheld.psi.cz/documents/FluorPen_Monitoring_Manual_02_2021.pdf .Salehi, B., M. Shivaprasad Shetty, N.V. Anil Kumar, J. Živković, D. Calina, A. Oana Docea, S. Emamzadeh-Yazdi, C. Sibel Kılıç, T. Goloshvili, S. Nicola, G. Pignata, F. Sharopov, M.D.M. Contreras, W.C. Cho, N. Natália, J. Sharifi-Rad. 2019. Veronica plants—drifting from farm to traditional healing, food application, and phytopharmacology. Molecules. 24(13):2454. https://doi.org/10.3390/molecules24132454
[Article] [PubMed] [PMC]Semchenko, M., M. Lepik, L. Götzenberger, K. Zobel. 2012. Positive effect of shade on plant growth: amelioration of stress or active regulation of growth rate? Journal of Ecology. 100(2):459-466. https://doi.org/10.1111/j.1365-2745.2011.01936.x
[Article]Shim, D., S.H. Jeon. 2022. Appropriate amount of nitrogen fertilizer for shading cultivation of tea tree seedlings. Horticultural Science and Technology. 40(6):643-653. https://doi.org/10.7235/HORT.20220058
[Article]Shin, H.T., M.H. Yi, J.S. Shin, B.C. Lee, J.W. Yoon. 2012. Distribution of rare plants-Ulsan, Busan, Yangsan. Journal of Korean Nature. 5(2):145-153. https://doi.org/10.7229/jkn.2012.5.2.145
[Article]Song, S.J., M.J. Jeong, S.Y. Kim, S.Y. Lee. 2020a. Phenotypic plasticity and ornamental quality of four Korean native Veronica taxa following different light intensity treatment. Flower Research Journal. 28(3):123-131. https://doi.org/10.11623/frj.2020.28.3.03
[Article]Song, S.J., M.J. Jeong, S.Y. Lee. 2020b;Ecophysiology of growth and flowering of four Korean native Veronica taxa in response to the photoperiod and cold treatment. In: III International Symposium on Germplasm of Ornamentals 1291; pp 205-214. https://doi.org/10.17660/ActaHortic.2020.1291.26 .
[Article]Song, S.J., U.S. Shin, H.J. Oh, S.Y. Kim, S.Y. Lee. 2019. Seed germination responses and interspecific variations to different incubation temperatures in eight Veronica species native to Korea. Horticultural Science and Technology. 37(1):20-31. https://doi.org/10.12972/kjhst.20190003
[Article]Spoustová, P., H. Synková, R. Valcke, N. Čeřovská. 2013. Chlorophyll a fluorescence as a tool for a study of the potato virus Y effects on photosynthesis of non-transgenic and transgenic Pssu-ipt tobacco. Photosynthetica. 51(2):191-201. https://doi.org/10.1007/s11099-013-0023-4
[Article]Srivastava, A., R.J. Strasser, G. Govindjee. 1999. Greening of peas: parallel measurements of 77 K emission spectra, OJIP chlorophyll a fluorescence, period four oscillation of the initial fluorescence level, delayed light emission, and P700. Photosynthetica. 36(13):365. https://doi.org/10.1023/A:1007199408689
[Article]Stirbet, A., G. Gavindjee. 2011. On the relation between the Kautsky effect (chlorophyll a fluorescence induction) and photosystem II: basics and applications of the OJIP fluorescence transient. Journal of Photochemistry and Photobiology B: Biology. 104(1–2):236-257. https://doi.org/10.1016/j.jphotobiol.2010.12.010
[Article] [PubMed]Strasser, R.J., A. Srivastava, M. Tsimilli-Michael. 2000. The fluorescence transient as a tool to characterize and screen photosynthetic samples. (pp. 445-483). In: Yunus M., Pathre U., Mohanty P., (Eds), Probing photosynthesis: Mechanisms, regulation and adaptation (pp. 445-483). London, UK: Taylor & Francis.Sultan, S.E. 1987. Evolutionary implications of phenotypic plasticity in plants. Evolutionary Biology. 21:127-178. https://doi.org/10.1007/978-1-4615-6986-2_7
[Article]Thach, L.B., A. Shapcott, S. Schmidt, C. Critchley. 2007. The OJIP fast fluorescence rise characterizes Graptophyllum species and their stress response. Photosynthesis Research. 94:423-436. https://doi.org/10.1007/s11120-007-9207-8
[Article] [PubMed]Weijschede, J., J. Martínkova, H. De Kroon, H. Huber. 2006. Shade avoidance in Trifolium repens: costs and benefits of plasticity in petiole length and leaf size. New Phytologist. 172(4):655-666. https://doi.org/10.1111/j.1469-8137.2006.01885.x
[Article] [PubMed]Yang, H.R., Y.J. Park, M.J. Kim, J.Y. Yeon, W.S. Kim. 2022. Growth responses of Korean endemic Hosta minor under sub-optimal artificial lighting. Horticultural Science and Technology. 40(3):286-295. https://doi.org/10.7235/HORT.20220027
[Article]Yoo, S.Y., K.C. Eom, S.H. Park, T.W. Kim. 2012. Possibility of drought stress indexing by chlorophyll fluorescence imaging technique in red pepper (Capsicum annuum L.). Korean Journal of Soil Science and Fertilizer. 45(5):676-682. https://doi.org/10.7745/KJSSF.2012.45.5.676
[Article]Zettl, A. 2023 April 12 Converting Colors. Converting Colors website Retrieved from https://convertingcolors.com .Živčák, M., M. Brestič, K. Olšovská, P. Slamka. 2008. Performance index as a sensitive indicator of water stress in Triticum aestivum L. Plant, Soil and Environment. 54(4):133-139. https://doi.org/10.17221/392-PSE
[Article]