3가지 목본성 정원식물의 맹아기에 필요한 적산온도 분석
Analysis of Accumulated Temperature According to Sprouting of 3 Garden Woody Plants
Article information
Abstract
The rapid urbanization led to construction of urban structures and buildings in uniform, damaging the urban landscape. One of the methods to enhance the urban landscape is expansion of green area. The problem is, however, there are much differences in blossoming seasons that are of same species and area, which is due to microclimate caused by many environmental factors and conditions. The differences in blossoming seasons deter harmonized arrangement for beautiful urban landscape. The objective of this study is to find the accumulated temperature for 80% of growth for sprouting so that blossoming season can be expected. To this end, Cornus officinalis Siebold & Zucc, Corylopsis gotoana var. coreana, and Forsythia koreana(Rehder) Nakai that are unrelated to accumulated temperature were collected in Suwon. Then, 30 flower buds were put in place in growth phases of 5, 10, 15, and 20°C, which repeated 4 times. The sprouting time of flower buds and temperature were checked every 2 days. It turned out that the higher the temperature was, the more facilitated the sprouting time was. However, excessive high temperature tended to deter the sprouting due to certain characteristics of plants. The spouting speed was the highest for cornus fruit. Fragrant winterhazel, and forsythia were the next. At temperature of 15°C, however, the sprouting speed of forsythia was higher than that of fragrant winterhazel temporarily. At 20°C, forsythia had higher sprouting speed than fragrant winterhazel, The accumulated temperature necessary for 80% of growth for sprouting was 169.3±7.7°C for C. officinalis Siebold & Zucc, 226.0±18.0°C for C. gotoana var. coreana, and 279.0±0.4°C for F. koreana(Rehder) Nakai.
I. 서론
산업혁명 이후, 인구의 도시집중과 공간 사회 · 경제적 측면에 서 도시적으로 변화해가는 현상 즉, 도시화의 가속이 붙기 시작하 였다. 도시화는 녹지면적을 감소시켰다. 이로 인하여 도시의 생활 권 녹지면적은 1인당 4.53m2로서 매우 저조한 실정이며(Ro et al., 2003) 획일적인 도시 구조물은 도시의 경관을 악화시켰다. 때문에 도시녹지에 대한 사회적 요구도는 점차 높아지고 있으며 걷고 싶은 거리, 꽃을 소재로 하는 도시 축제 등과 연계되어 도시녹지의 중요 성은 증가하고 있다(Kim et al., 2013).
이러한 도시 경관을 개선하기 위해서는 도심지 녹지 면적의 확 대가 필요하다. 하지만 도시는 공간별 다양한 환경조건이 구성되어 다양한 미기후가 만들어진다. 이로 인하여 동일 종 · 지역이라도 고 도, 사면, 가지위치 등에 따라 개화기의 차이가 발생한다(Jo and Ahn, 2008). 이러한 문제는 도시 경관을 고려한 녹지 조성 시 식물 사용의 어려움을 준다.
식물계절학은 연중 식물 생장에 따른 현상의 차이로 생기는 징 후와 지속에 대한 연구로서 온도, 습도, 일조시간 등의 기후조건의 변화에 식물이 어떠한 적응방식을 보여주는가를 나타낸다(Fenner, 1998). 식물의 개화나 개엽에 영향을 주는 환경인자는 기후, 토양 의 수분, 일장, 식물체내의 수분포텐셜 등 인데 특히 겨울 및 봄철의 온도가 가장 민감한 요인이다(Lee and Ho, 2003). 적산온도의 개 념은 1730년대 프랑스 과학자인 Rene A. F. de Reaumur에 의해 소개되었으며 적산온도를 이용하여 식물 또는 곤충 등 생물의 성장 단계를 예측 할 수 있다(McMaster and Wilhelm, 1997). 그리고 이러한 식물활동과 온도와의 관계는 기온과 시간 개념으로 확대되 어(Lindsey and Netman, 1956) 일적산온도, 온량지수, 연일지수 등이 제시된 바 있다(Yim et al., 1983; Yim, 1986; Yim, 1987).
관련 연구사례로는 한국의 온도기후와 생물의 계절변화에서는 연일지수(year day index)를 이용하여 적산온도를 계산하였고 (Yim et al., 1983), 생육초기 수 종 목본식물의 화력학적 특성에서 는 일 적산온도(Tn)를 이용하여 관목류의 개화 시 적산온도를 계산 하였다. 그리고 수 종 목본식물의 개화시기와 개화기간에 관한 연 구에서는 연일지수(year day index)와 일 적산온도(Tn) 2가지 방 법으로 계산 후 적산온도량과 개화기의 상관관계를 분석하였다. 또 한 Rubus의 만개일을 예측하기 위해 linear와 curvilinear 모델을 이용하여 2가지 품종의 주요온도 및 적산온도(growing degree hour)의 설정한 바 있으며(Black and Frisby, 2008), GDD=[(Tmax+Tmin)/2] - Tbase 공식을 이용하여 Hedysarum boreale 성 장 단계별로 heat unit을 계산하였다(Peel et al., 2011). 그 외에도 발육속도모델을 이용한 복숭아 ‘유명’의 개화기 예측에 관한 연구 (Yun et al., 2012), 한반도 식물계절과 기후에 관한 연구(Lee, 2011), 점봉산에서 미기후 변화에 따른 목본식물의 봄철 식물계절 학 연구(Lee, 2012)이 연구 되었다.
본 연구에서는 봄철에 개화하는 세 가지 목본성 조경식물의 맹 아율 80%까지 필요한 적산온도를 구명하고자 한다.
II. 연구방법
중부지역에서 자생하는 목본식물 중 봄철에 개화하는 3가지 조 경식물을 대상으로 2014년 1월 14일부터 2월 28일까지 진행하였 다. 시험재료는 1월 14일에 수원 지역에서 수집하였으며 대상식물 은 산수유, 히어리, 개나리이다. 수집된 재료는 수집 즉시 꽃눈이 붙 어있는 상태로 15cm 길이로 절취되었으며 증류수를 넣은 플라스 크병에 꽂아 꽃눈 30개씩 4반복으로 5, 10, 15, 20°C 유지되는 식 물생장상에 배치하였다(Fig. 1). 그리고 2일 주기로 맹아율을 조사 하였다. 데이터로고(Watch dog, Spectrum, USA)를 이용하여 온 도데이터를 수집하였으며 측정은 1시간 간격으로 0.1°C 단위로 측 정하도록 조작하였다. 적산온도를 계산하는 방법은 개화 시 까지 식물생장상의 일평균온도를 적산하여 계산하였다. 그리고 계산된 적산온도를 실증하기 위하여 2013년 수원지역의 정원식물 개화기 현장조사 기록을 활용하였으며 적산온도 계산방법은 일평균기온 이 0°C 이상인 온도를 누적한 연일지수(year day index; Yim, 1987)를 활용하였다. 연일지수의 계산식은 다음과 같다.
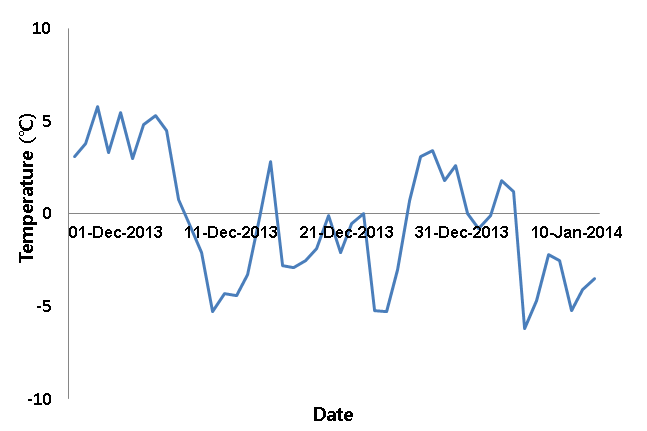
Daily mean temperature in Suwon from December 2013 to January 2014.
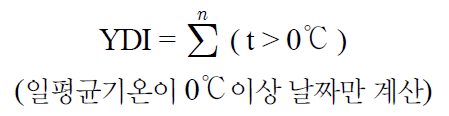
데이터는 SPSS(varsion. 21)로 통계처리를 하였으며 mean±S.D. 로 표시하였다. 자료에 대한 유의성 검정은 일변량 분산분석 (ANOVA)에 의하였으며 p-value가 0.05미만의 경우를 유의한 것 으로 하였다.
III. 결과 및 고찰
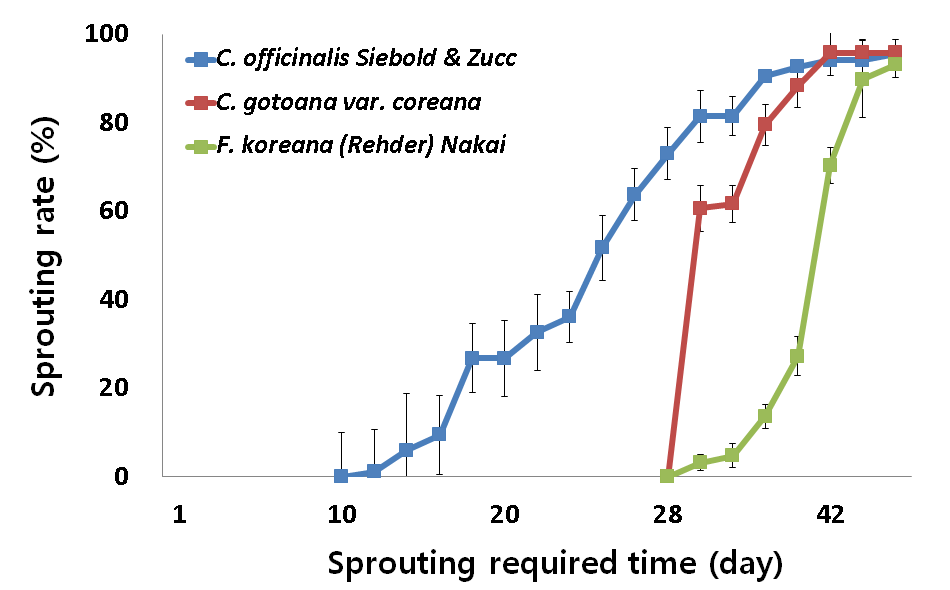
3 가지 조경식물 산수유, 히어리, 개나리의 꽃눈을 5°C, 10°C, 15°C, 20°C 수준별로 처리한 결과는 Fig. 2~5와 같다. 5°C, 10°C, 15°C, 20°C에서 3가지 식물은 모두 맹아율 80% 이상 정상적인 맹 아 활동을 나타내었다. 하지만 산수유, 히어리, 개나리는 온도수준 에 따라 다른 맹아율과 맹아 속도를 나타내었다.
5°C Temperature Conditions according to C. officinalis Siebold & Zucc, C. gotoana var. coreana, F. koreana(Rehder) Nakai of sprouting rate. All data are means±SD of four replication.

20°C Temperature Conditions according to C. officinalis Siebold & Zucc, C. gotoana var. coreana, F. koreana(Rehder) Nakai of sprouting rate. All data are means±SD of four replication.

10°C Temperature Conditions according to C. officinalis Siebold & Zucc, C. gotoana var. coreana, F. koreana(Rehder) Nakai of sprouting rate. All data are means±SD of four replication.
맹아 속도는 그래프의 기울기가 급하게 나타난 고온에서 빠르게 나타났으며 식물별 개화속도는 산수유, 히어리, 개나리 순이었다. 하지만 15°C 수준에서는 16일차부터 개나리가 히어리보다 맹아속 도가 빠르게 나타났으며(Fig. 4) 20°C 수준에서는 대체적으로 개 나리가 히어리보다 맹아속도가 빠르게 나타났다(Fig. 5).
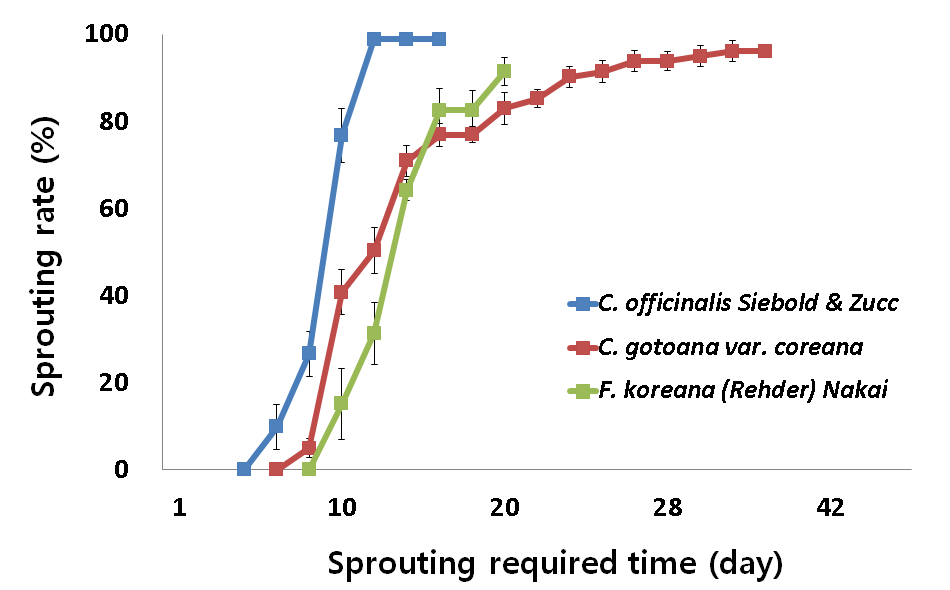
15°C Temperature Conditions according to C. officinalis Siebold & Zucc, C. gotoana var. coreana, F. koreana(Rehder) Nakai of sprouting rate. All data are means±SD of four replication.
산수유, 히어리, 개나리는 온도 수준에 따라 맹아 80% 수준까지 다른 적산온도 요구량을 나타내었다. 맹아율 80% 수준까지 산수 유는 5, 10, 15°C 수준에서는 적산온도 요구량의 차이가 없었으나 5, 10, 15°C와 20°C 수준 간 에는 차이가 발생하였다. 20°C 수준에 서는 고온에 의해 맹아기가 촉진된 것으로 보인다. 히어리는 5, 10°C 수준에서는 적산온도 요구량의 차이가 없었으나 5와 10°C, 15°C, 20°C 수준 간 에는 차이가 발생하였다. 15°C 수준에서는 고 온에 의해 맹아기가 촉진된 것으로 보이나 20°C 수준에서는 고온 에 의해 맹아기가 억제된 것으로 보인다. 개나리는 5, 10°C와 15, 20°C 수준에는 적산온도 요구량의 차이가 없었으나 5, 10°C와 15, 20°C 수준 간에는 차이가 발생하였다. 15, 20°C 수준에서는 고온 에 의해 맹아기가 촉진된 것으로 보인다(Table 1).
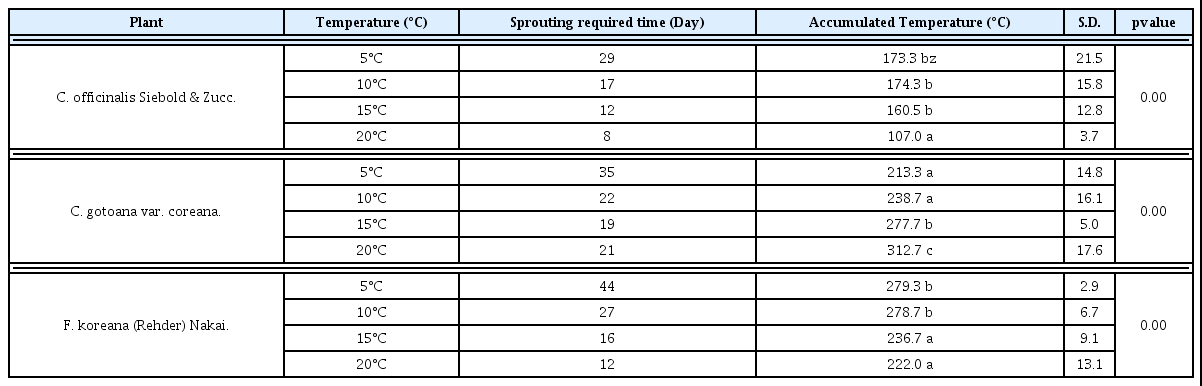
Accumulated Temperature and duration ccording to sprouting rate of 80% of 3 garden woody Plants.
대체적으로 처리 온도가 고온일수록 맹아기를 촉진 하였지만 식 물의 특성에 따라 일정 수준 이상의 고온은 식물의 맹아기를 억제 하였으며 그 원인은 최고온도 수준까지 올라가서 맹아 활동에 관련 된 요인들의 활성도가 약화되어 식물의 생장을 감소시키기 때문인 것으로 예상된다(Mwale et al., 1994; Shafii and Price, 2001). 연 구 결과로 인하여 맹아기에 영향을 주는 주요요인을 특정 지을 수 없지만 식물에 따라 맹아의 최적온도가 다른 점에서 단일요인보다 는 복합요인일 것이라 생각된다. 관여 요인 및 온도에 의한 요인의 활성화 정도는 종 특이성이 반영된 것이라 예상되며 요인들의 특성 에 따라 주요온도가 다르다. 때문에 온도수준에 따라 식물별로 다 른 맹아 활성도를 보인 것으로 생각된다. 이것은 식물의 생리현상 에 영향을 주는 호흡, 수분흡수, 물질전류, 호르몬의 생리작용의 주 요온도는 각각 다르다(Chae et al., 2006)라는 내용과 유사하다.
이상 결과와 봄철 일 평균기온 고려 시 맹아 80% 수준에 필요한 적산온도는 산수유 169.3±7.7°C, 히어리 226.0±18.0°C, 개나리 279.0±0.4°C로 나타났다. 연구결과를 검증하기 위하여 2013년 수 원 지역의 노지에 식재된 산수유, 히어리, 개나리의 맹아 80%까지 개화일과 적산온도를 자료를 사용하였다. 2013년 연구자료에서 계 산한 적산온도는 산수유 196.2°C, 히어리 241.2°C, 개나리 327.6°C 이었으며 두 자료를 비교 시 적산온도 편차는 산수유 ±18.97°C, 히 어리 ±26.73°C, 개나리 ±71.84°C로 나타나서 본 연구결과와 유사 하였다. 그리고 개나리는 서울시 남산에서 자생지에서 개화 시 까 지 적산온도가 2005년 298.2°C, 2006년 261.2°C(Lee, 2007)라 고 주장하여 본 연구결과인 279.0±0.4°C와 유사한 수준을 나타내 었다.
적산온도를 이용한 식물 생장력 예측 방법 중 Lindsey & Newman (1956)이 제안한 적산온도 계산법은 6.1°C를 생리적 활성화 임계 온도로 보아 그 이하의 온도를 잘라낸 온도를 적산하여 온량을 나 타내는 지수로 삼았다. 하지만 본 시험에서는 5°C 수준에서도 3종 의 식물의 맹아가 정상적으로 진행되었다. 때문에 봄철 맹아기를 거쳐 개화하는 식물의 최저 생장온도는 5°C 보다 더 낮을 것이라 생 각된다. 이는 대부분의 식물종의 생리적 활성화 임계온도 즉 최저 온도는 종에 따라 0~5°C(Monteith, 1981)이라는 주장과 유사하 며 5°C이하의 온도를 제한 후 온도를 적산하는 일적산온도법이 온 도가 높은 남해안 등지에서 이른 봄에 개화하는 식물들의 개화 현 상을 설명하는데 부적당하다고 주장한 사례와 유사하다(Yim, 1987). 식물의 최저온도에 관하여 다양한 의견이 존재하는 이유는 식물종에 따라 최저온도가 다르며 이는 꽃눈 세포의 유리 당 및 호 르몬의 차이에 의해 꽃눈 세포의 빙점이 다르기 때문이다(kim et al., 2009). 본 연구에서는 생리적 활성화 임계온도를 0°C 설정한 연일지수를 활용하여 3가지 목본성 정원식물의 맹아 80%수준까 지 필요한 적산온도를 계산하였다. 하지만 보다 정확한 적산온도의 계산을 위해서는 식물 종에 따른 최저생장온도 설정에 관한 연구가 선행되어야 할 것이다.
IV. 적요
급속한 도시화로 인하여 도시 내부에 획일적인 도시 구조물이 건설되었다. 이것은 도시의 경관을 악화시켰다. 이러한 도시 경관 을 개선하기 위한 방법은 도심지 녹지 면적의 확대이다. 하지만 도 시 공간은 다양한 환경조건에 의해 미기후가 만들어지고 이로 인하 여 동일 종 · 지역이라도 개화기의 차이가 발생한다. 이러한 식물계 절의 차이는 도시 경관을 고려한 녹지 조성 시 식물 사용의 어려움 을 준다. 때문에 본 연구에서는 식물계절 예측을 위하여 맹아 80% 수준까지 필요한 적산온도를 구명하였다. 연구 결과 대체적으로 처 리 온도가 고온일수록 맹아기를 촉진하였다. 하지만 식물의 특성에 따라 일정 수준 이상의 고온은 식물의 맹아기를 억제하는 현상이 나타났다. 식물별 개화속도는 산수유, 히어리, 개나리 순이었다. 하 지만 15°C 수준에서는 일시적으로 개나리가 히어리보다 맹아속도 가 빠르게 나타났으며 20°C에서는 개나리가 히어리보다 맹아속도가 빠르게 나타났다. 그리고 맹아율 80% 수준에 필요한 적산온도는 산수유 169.3±7.7°C, 히어리 226.0±18.0°C, 개나리 279.0±0.4°C 나타났다.