Where Are Landscape Designers' Spatial Abilities in the Brain? An fMRI Study
Article information

Abstract
Background and objective
To effectively understand and communicate their work, landscape designers should possess excellent spatial abilities. Neurological methods have confirmed that activation of the occipital lobe, parietal cortex, and prefrontal cortex affect the judgment of space; however, few studies have measured spatial abilities in landscape design. This study aimed to identify the potential role of various brain regions during spatial interpretation processes by landscape designers, particularly the effect of stimulating the frontal lobe on enhancing design capabilities.
Methods
This study tested the spatial abilities of landscape designers when transforming a planar drawing into a sectional drawing and the brain regions activated in this process. The subjects were asked to identify the correct option when matching given section lines in a planar drawing. The correct answer rate and response time were used to score brain activation during spatial task processes. A total of 16 valid subjects were divided into high- and low-accuracy groups according to the correct answer rate.
Results
The results for the high-accuracy group showed that the left inferior frontal gyrus was activated during spatial design tasks. In contrast, the findings for the low-accuracy group revealed that the left middle occipital gyrus was activated for processing visual information.
Conclusion
The findings suggest that the frontal lobe plays a role in allowing landscape designers to make planar to cross-sectional inferences via mental rotations and categorical spatial relations. The findings offer implications for landscape designers in stimulating the frontal lobe and enhancing their design capabilities.
Introduction
Spatial ability holds significant relevance for the general public and landscape designers in particular, as it governs our sense of direction and spatial awareness. Goel (2014) introduced the Frontal Lobe Lateralization Hypothesis (FLLH), which postulates that the left and right prefrontal cortices (PFC) fulfill distinct roles in the design process. In addition, the emerging field of neurocognition in design, as highlighted by Balters et al. (2023), offers insights into the interplay between spatial relations, orientations, visualizations, and brain activations, which are integral components of the cognitive processes employed by landscape designers. Notably, Tsai et al. (2021) identified a link between graphic design thinking in landscape architects and the left prefrontal cortex, while the refinement of design drafts was associated with the activation of the left middle frontal gyrus.
Furthermore, investigations such as that by Suh and Cho (2020) have explored the positive correlation between visual-spatial abilities, mental rotation, and creativity in interior design. Likewise, Lopez et al. (2020) conducted independent research on the unconditional and coordinated relations in map-based tasks. A more direct and objective understanding of the relationship between landscape design and individuals' spatial skills can be achieved by capturing brain activation regions instead of relying solely on self-reports of behavior or psychological responses. Hence, employing functional magnetic resonance imaging (fMRI) to analyze brain images has become crucial in investigating the connection between cerebral reaction zones and the spatial interpretation abilities of landscape designers.
Literature review of spatial abilities in landscape design
Dimensions of spatial ability
According to one theory of multiple intelligence, spatial abilities permit the recognition and manipulation of 2D graphics in a 3D space (Gardner, 2011). Three factors in spatial ability purposed by Thurstone (1950) and Smith (1964) describes those terms more deeply, i.e., (1) mental rotation, (2) spatial visualization, and (3) spatial perception. Moreover, Pellegrino and Hunt (1991) highlighted another factor, (4) dynamic spatial ability, which is similar to Lohman's (1979) reference to mental spatial transformations. Mental rotation refers to the recognition of an object's directional change (Lohman, 1979). Thurstone (1938) identified seven factors, called primary mental abilities, associated with memory, reasoning, and spatial visualization. One of these factors, "space," was related to spatial or visual graphics. McGee (1979) proposed two factors: spatial visualization and spatial orientation. Spatial visualization is the ability to recognize an object's partial or motional change and involves the mental manipulation, rotation, and inversion of stimuli. Spatial orientation involves correctly recognizing changes in the vertical or horizontal dimensions of stimuli in two to three dimensions, a concept similar to spatial perception, which relates spatial orientation to one's body (Mohler, 2009). Dynamic spatial ability is involved in the process of motion or relative motion.
Spatial ability and the related mechanism in brain activation
Landscape designers rely on spatial abilities involving imagery and categorical and coordinate spatial relations to accurately analyze and rotate a 2D planar map into 3D sections. For instance, when designers view actual images, the images are imprinted on their mind, which in turn enables them to convert the spatial orientation of landscape elements and categorize and coordinate them fluently. These abilities, which deal with visual imagery and visual perception, are involved in two different neural mechanisms-"top-down voluntary imagery signals" and "bottom-up perception"-that have important functions concerning spatial ability (see Fig. 1).
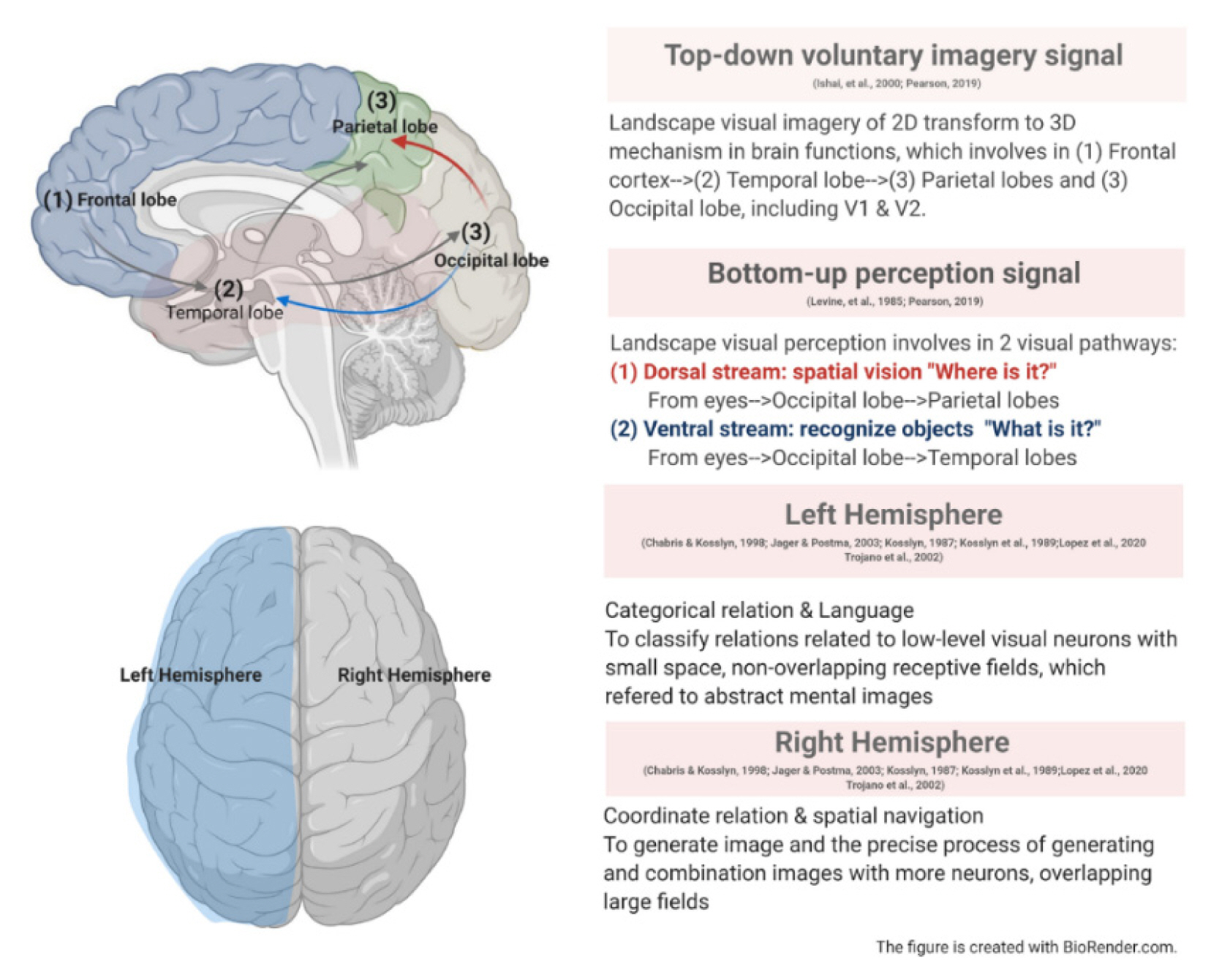
Spatial ability and related brain mechanisms involved in landscape design.
The top-down voluntary imagery signal mechanism involves high-level areas of the brain, including the frontal cortex, which triggers a backwards route to stimulate related information in the temporal lobe and parietal lobe, ultimately involving the primary visual cortex to form visual images (Ishai et al., 2000; Pearson et al., 2019). It is argued that mental imagery is associated with visual areas, such as V1 and V2 for space and orientation, and can also be related to the default mode network in a resting state (Dijkstra et al., 2017; Naselaris et al., 2015; Pearson et al., 2019). The top-down voluntary imagery signal also overlaps with the visual imagery and visual perception areas (Pearson et al., 2019) that induce spatial transformations related to another brain mechanism: bottom-up perception. The bottom-up perception mechanism operates from the eyes to the primary visual cortex (occipital lobe), which is associated with two visual pathways responsible for processing spatial information from objects: (1) the dorsal stream, which engages in spatial vision, and (2) the ventral stream, which recognizes objects (Levine et al., 1985) and transmits "positions" and "images" to the parietal lobe, temporal lobe, and frontal lobe. In one graphics-based study, it was found that the left and right superior parietal cortices, bilateral prefrontal and occipital-temporal cortices, and insula and basal ganglia regions were all involved in visuo-spatial reasoning among children and teenagers (Eslinger et al., 2009) (see Fig. 2). Another study showed that the parietal lobe is related to drawing and that parietal activations are higher in the left than the right side of the brain (Makuuchi et al., 2003) (see Fig. 2). Furthermore, the authors stated that two visual pathways are involved in the drawing process: the dorsal stream, which is stimulated by visual information to capture an object's frame and related spatial structure, after which the parietal lobe stimulates the dorsal premotor area (BA 6) and ventral premotor area (BA 44); and the ventral stream, which recognizes objects via the inferior temporal sulcus (BA 37) (Makuuchi et al., 2003).

Interactions between the frontal lobe and parietal lobe explain the relationship between brain function and spatial design.
Two spatial relations-categorical relations and coordinate relations-are important skills in landscape design (see Fig. 1). Categorical relations refer to the capturing of elements/properties in space without pinpointing an accurate location; in other words, they represent the spatial relative position of each element, such as right or left. Coordinate relations, on the other hand, compute the precise position in the space (Jager and Postma, 2003; Trojano et al., 2002). Kosslyn (1987) derived theories on spatial relationships and orientations of the parietal lobe from categorical relations and coordinate relations. How do the theories on spatial relationships and orientations mentioned in the preceding sentence relate, in a progressive or relational sense, to the language capacity of the brain's left hemisphere mentioned in the following sentence? As well, the left hemisphere of the brain involves language, providing the ability to classify relations of low-level visual neurons with small-space, non-overlapping receptive fields, whereas the right hemisphere involves more neurons with overlapping and large fields that dominate spatial navigation in coordinate relations (Chabris and Kosslyn, 1998; Jager and Postma, 2003; Kosslyn, 1987; Kosslyn et al., 1989; Trojano et al., 2002). One study, which employed an imaginative "coordinate" mental clock test, asked subjects to imagine a mental clock and use it to categorize the same direction of a pair of numbers, which were presented in the mental clock as a "categorical task." Both "coordinate" and "categorical" tasks were used as experimental stimuli. The study found that visual perception was related to spatial imagery and categorical and coordinate spatial judgments, which are activated in the superior parietal lobule and also in the parietal and prefrontal areas. That is, the parietal lobe refers to spatial coding and graphic spatial mind thinking, while the superior parietal lobule processes the spatial transformation area (Trojano et al., 2002) (see Fig. 2). This implies that the coordinate relation is activated in the right prefrontal cortex in the right hemisphere, which is related to the generation of images and the precise process of generating and combining images with higher processing loading on working memory in spatial tasks, whereas the categorical relation is associated with the whole picture and with abstract mental images in the left hemisphere (Trojano et al., 2002). Further research, which asked students to measure and map campus landmarks, lent support to the theory of hemispheric lateralization by showing that categorical and coordinate relations occurred in separate brain regions (Lopez et al., 2020) (see Fig. 1). Above all, spatial abilities in landscape design and planar mapping involve complex cognitive processes, which must use mental rotation, spatial visualization, and spatial perception to rotate a planar drawing into a sectional drawing to communicate with others.
Spatial ability plays a crucial role in landscape design, encompassing various cognitive processes involved in spatial reasoning, such as translating plans, cross-sections, and perspectives. These processes engage critical regions of the brain, including the frontal lobe, which is responsible for decision-making, problem-solving, and attention; the prefrontal cortex, which governs judgment, reasoning, and planning; and the parietal lobe, which processes sensory information, facilitates visual-spatial processing, and aids in spatial recognition and short-term memory. Transforming plans into cross-sections requires spatial reasoning skills akin to identifying and locating various points on a geographic coordinate system. Mental rotation and the ability to comprehend categorical and coordinate relations are utilized to understand relative positions and precise distances on sketchy maps, as demonstrated by Lopez et al. (2020). Moreover, transitive inference, an ordered reasoning test, has been found to activate the frontal and parietal lobes, exhibiting partially spatial-like operations and mental rotation, as reported by Acuna et al. (2002) (see Fig. 2).
Given these insights, the present study posited that spatial ability is intricately connected to reasoning, planning, and visual-spatial cognition in landscape design. An exploratory approach utilizing fMRI technology was employed to examine this relationship. The study aimed to identify specific brain regions associated with high spatial ability among landscape designers, thereby contributing to the existing knowledge base in landscape design.
Research Methods
The fMRI method was suitable for achieving the research goal because it allowed the non-invasive measurement of brain activity in response to various stimuli or tasks. Researchers can determine which brain regions are involved in the experimental processes and how they are connected by examining the patterns of brain activation involved in converting a planar drawing to a sectional drawing, which, in the current case, helped in the evaluation of the research hypothesis (Fig. 3). Therefore, an fMRI experiment was employed to directly investigate the neural basis of spatial interpretation among landscape designers, ultimately providing valuable information for other researchers in this area.
The research hypothesis in the study.
The experiment was separated into two parts. First, experimental subjects were selected by inviting 12 individuals with experience in landscape design (having completed at least four years in landscape design or related landscape courses) and 12 novices (those with no background in landscape design or a related design field) to take a pre-test for selecting the experimental materials. By doing so, the study could control for the quality of the difficult sequence and to select the dependability of experimental materials employed in the formal test. Next, for the fMRI formal test, 32 individuals with experience in landscape design and novices were invited to join. Each subject gave signed informed consent, which was approved by the Research Ethics Committee [approval number 201503HM004] (Fig. 4). The inclusion/exclusion criteria for the fMRI experiment were as follows:
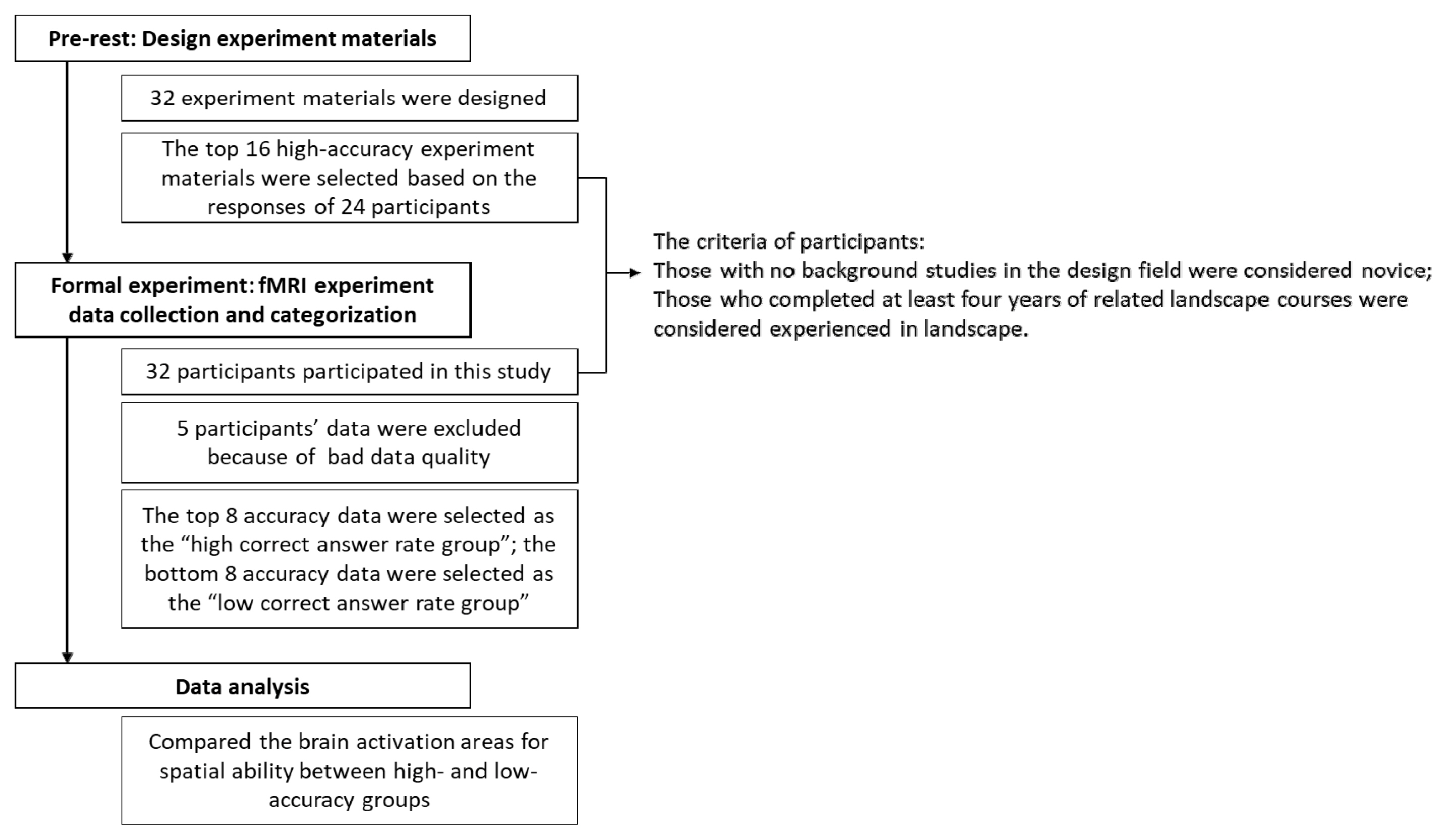
The workflow of the study.
(1) Be between the age of 20 and 30 and either be currently enrolled in a university or possess a university degree. Additionally, have completed at least four years of coursework in landscape studies, which was considered as having experience in landscape design.
(2) Speak Chinese as a native language, with proficiency in comprehending Chinese text.
(3) Be naturally or habitually right-handed.
(4) Must not have metal dentures, fixed metal braces, or orthodontic appliances implanted in the oral cavity.
(5) Must not have any implanted metal objects, such as steel nails, cardiac pacemakers, artificial heart valves, cochlear implants, insulin injectors, implanted drug injectors, internal electrodes, stimulators, nerve stimulators, or vascular stents.
(6) Have no history of emotional or mental illnesses, brain injuries, cardiovascular diseases, epilepsy, or similar conditions.
(7) Must not have undergone any heart or brain surgeries in the past.
(8) If myopic, the degree of myopia must not have exceeded 900 degrees, and vision must have returned to normal after correction; the wearing of contact lenses was permissible.
(9) Must not have been currently experiencing sinusitis or common cold symptoms.
(10) Must not have been pregnant at the time of the experiment.
(11) Must not have been currently taking medications or habitually consuming alcohol.
(12) Must not have been claustrophobic.
(13) Must not have had a body weight exceeding 100 kg.
The data collection instruments used in the study were assessed by the Magnetic Resonance Imaging Laboratory with a Siemens Magnetom Prisma MRI at a magnetic field strength of 3T and equipped with a head-sized ramp and headphones for protecting the load noice at the XXX (for double-blind review process). A Siemens Prisma 3T MRI scanner was used with the following specifications: TR = 3000 ms, TE = 30 ms, FOV = 192 mm, number of slices = 45, and voxel size = 3 × 3 × 3 mm. The scan time was 10 min, which was set to "interleaved" from ventral to bottom-up. Goggles were placed separately on the head (VisuaStim XGA, Resonance Technology Inc., CA, USA) for viewing the experimental materials.
Pre-test
A specially designed "planar/cross-section space test" with three objects-trees, houses, and bushes-was used in the experiment. The layouts combined and arranged the three objects into 32 plans to test the subjects' spatial awareness of the landscape (see one of the examples in Fig. 5 for an illustration). First, the subjects saw a planar view on the left-hand side of the screen, with a sectional line A-A'. Simultaneously, the right-hand side of the screen showed the four sectional options, of which only one was correct, and the subjects had 30 s to answer. In the "pre-test experiment," 24 subjects, including both novices and those experienced in landscape, were asked to distribute the 32 experimental questions evenly. Then, Excel was used to rank correct answers (ACA) and E-prime accuracy to record their reaction times and accurate answer rates for each item. As a result, the final 16 experimental questions were developed with a high ACA, in which the spatial ability part of the brain was utilized. The typical response time was 27.66 s. As a result, each experimental question in the study had a 30 s time limit.

Two examples, (a) (b), of pre-tests for testing spatial ability in the brain.
Data collection procedure
Subjects were tested individually in the fMRI laboratory via the following process. First, the subjects were asked to sign the informed consent form, which explained the experimental process, ensured that the health requirements were met, and informed them that no harm would be caused by the fMRI experiment. Second, the subjects were informed that a security check would be used to calibrate the instrument for about 20 min and that they would be given earplugs and earmuffs to protect them from the loud sound made by the instrument. Additionally, they were informed that they would be given googles, a reaction button box to hold with both hands, and a safety bell to wear on their stomach in the event of an emergency. Finally, before the experiment, a structural scan of the whole brain was conducted, lasting 10–15 min, after which the spatial recognition tests, including the experimental description, sample examination question, and two sets of experiments, which could be completed in 15 min, commenced. After completing the fMRI experiment, the operator removed all of the experimental equipment, and the subjects were asked to go to the waiting room to receive a gift card. The neurophysiological scan settings were as follows: (1) the structural scan: (T1) sets TE = 46 ms, TR = 3000 ms, flip angle = 30 degrees, number of scans per experiment=45 slices, voxel size = 256 × 256 pixels (0.9375 × 0.9375 mm) in each x-y image, scanning thickness = 3.7 mm; (2) the functional scan: (T2) sets TE = 35 ms, TR = 3000 ms, flip angle = 90 degrees, voxel size = 64 × 64 pixels (3.75 × 3.75 mm) in each x-y image, scanning thickness=3.7 mm.
Formal experiment
In the block design experiment, two runs were performed, each consisting of eight blocks of functional scans. Before the experiment, eight slides were shown for introduction and practice. Next, a cross was displayed at the center of the screen for 20 s to attract the subjects' visual attention. Each experimental stimulus lasted for 30 s and was repeated eight times. The total scan time was 392 s. The procedure for the second run was the same as that for the first run, except that the introduction and practice slides were not shown (Fig. 6).
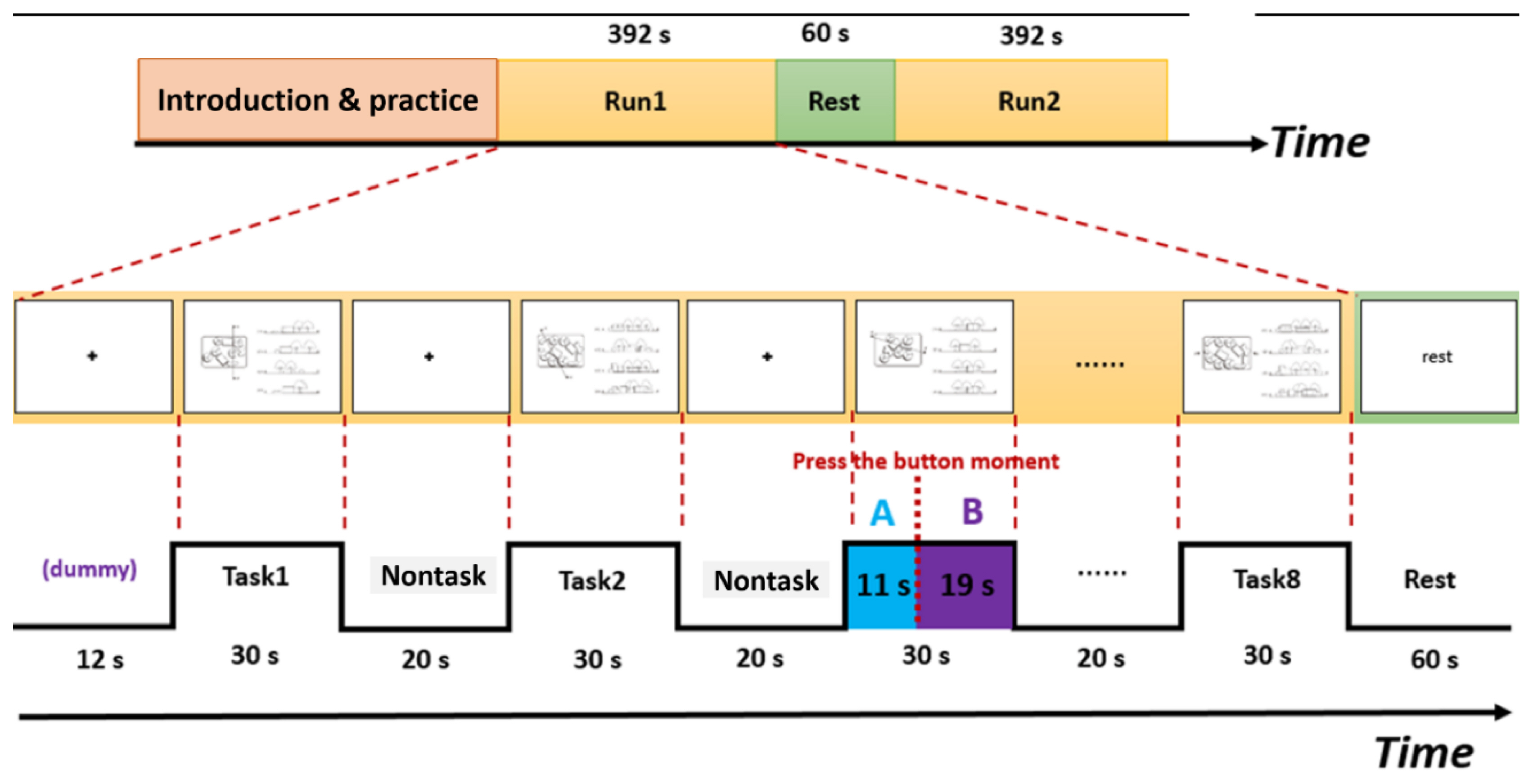
The block design of the fMRI experiment with two runs, each consisting of eight blocks of functional scans and an example of the A status minus B status, corresponding to brain activation related to spatial processes minus brain activation related to mind-wandering, which was defined as spatial abilities associated with landscape design in this study.
Data analysis
For the action response analysis, test response times and test answer rates were recorded in the E-DataAid File of the E-prime system, and E-merge was used to process the file data. The data were then imported into Excel for the purpose of performing descriptive statistics. For brain activation, the data processing comprised two parts: (1) preprocessing, correction, and alignment of each subject's brain image with standard brain coordinates, and (2) general linear model analysis, which involved examining the statistical significance of the brain activation area.
In the formal test, 32 subjects participated, each of whom answered 16 questions. For those whose correct response rate was under 75%, the data were excluded; therefore, two outliers were removed. In order to obtain accurate brain region images, one subject with an abnormal brain structure and one whose head movements affected the alignment of the brain image were not included for further analysis. Furthermore, to detect the connection between the press button and the hemisphere motion area, a sanity check was conducted (Fig. 7). During the sanity check, when subjects pressed the left-hand-side button, a response should be seen in the right hemisphere motor area, and vice versa. The brain image showed right brain motor cortex activation when we analyzed the period of using the left button minus the right button. If not, this data is not qualified to further analysis. One outlier was removed in this stage.

Reaction for left- and right-hand motion regions in the motion sanity check.
A total of 27 pieces of data were used to answer the questions concerning the relationships between the spatial ability of subjects experienced in landscape and novices. The results showed that the average accuracy correct answer rate (ACA) of the former was 0.84, while that of novices was 0.80, indicating no significant difference between the two groups. The data from the high correct answer rate (0.94) to the low correct answer rate (0.75), which included 11 subjects with an average 0.81 ACA rate, were rearranged. Therefore, data with an accuracy > 88% were classified into the high-accuracy group and included eight subjects, whereas data with an error rate > 25% were classified into the low-accuracy group and also included eight subjects, including those experienced in landscape and novices (see the description of the subjects in Table 1). Thus, in total, data from 16 subjects in the high- and low-accuracy groups were ultimately used to compare their brain activation areas.

The description of 16 subjects and their rate of correct answers
First, 1st-level analysis was used for individual brain area response. Then, 2nd-level group analysis was employed to determine the statistical significance of brain activation. In the 1st-level analysis, a general linear model was developed for each subject to analyze the spatial processing period. The total response time for each spatial ability question was 30 s. At the time the participant pressed the response button, the reaction time of this question was recorded. For example, if a subject took 11 s to view a question and press the response button, it was recorded as A status, while the remaining 19 s was considered as viewing the question plus mind-wandering, which was recorded as B status (see Fig. 6). A status included vision, pre-response spatial processing, and motor signals; B status included vision, post-response mind-wandering, and motor signals. A status minus B status (A–B)-namely, brain activation related to spatial processes minus brain activation related to mind-wandering-was defined as brain activations associated with spatial abilities in landscape design, which involved a state that was fully focused on and recognized the landscape spatial relationship in plan and cross-section. Therefore, the A-B status was used for analyzing brain activation in landscape spatial processes. After this, the 2nd-level group analysis was calculated the parameter estimates and contrast estimates from each subjects' 1st-level analyses data.
Results and Discussion
The brain execution spatial ability in landscape design was set as A status minus B states in each subject. The high-accuracy and low-accuracy groups were then tested separately using a one-sample t-test in SPM8 to understand the statistically significant brain region activity in each group. The results showed the different brain areas involved.
Results of brain activation areas for spatial ability in high- and low-accuracy groups
The brain activation area for spatial ability in the high-accuracy group is located in the inferior frontal gyrus [(−52, 28, 28) t = 22.50, pFWE-correct = 0.008)]. This finding suggests that the triangular part, which is part of the inferior frontal gyrus, involves a high level of cognition and was responsible for good spatial ability. On the other hand, the brain activation area for spatial ability in the low-accuracy group is located in the middle occipital gyrus [(−36, −84, 14) t = 24.17, pFWE-correct = 0.005)]. This is the area responsible for processing visual information. The results showed that people with poor spatial ability must expend more effort to process visual stimuli (Table 2 and Fig. 8).

Brain activation in high- and low-accuracy groups
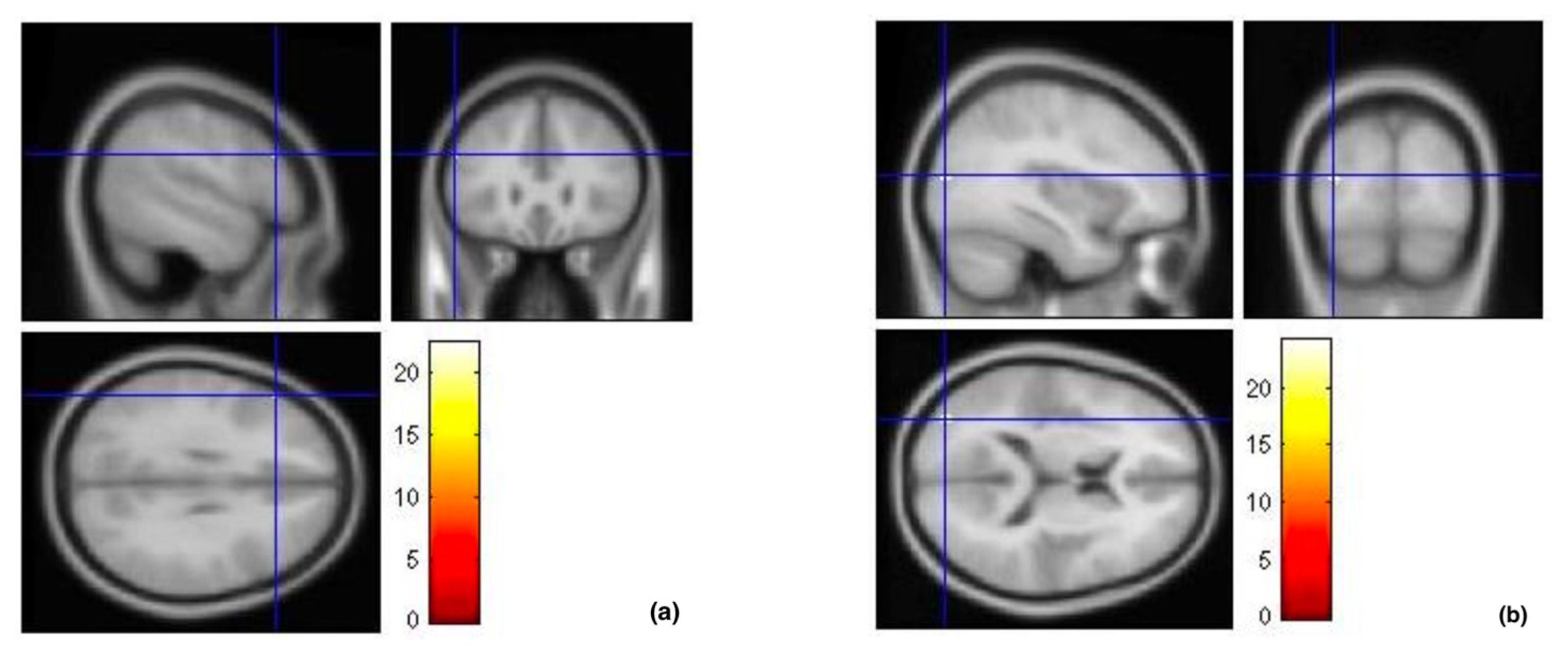
The cross-sections shown in these images indicate the brain activation areas in each group: (a) left inferior frontal gyrus was activated in the high-accuracy group, and (b) left middle occipital gyrus was activated in the low-accuracy group.
Discussion of brain activation in landscape design
This study constituted an initial effort to understand spatial ability at a design scale. From the literature review, it was inferred that landscape design involves the mechanisms of visual spatial imagery and the ability to coordinate and categorize spatial judgment. The 2D planar-to-3D cross-section findings of this study provide a knowledge base for designer-training pedagogy. Given the exploratory nature of the study, its findings also have implications for landscape design and, based on this, it should be treated with caution. In landscape design, spatial abilities, spatial visualization, and mental rotation involve a higher-level cognitive executive function, one related to the ability to understand plans (2D) and then transform them into cross-sections (3D). By using fMRI, the study provided explanations of the high spatial ability in brain activation while conducting graphic mental rotations in landscaping.
First, among the subjects, who were college students, there was no significant difference between those experienced in landscape and novices; therefore, the groups were rearranged into high- and low-accuracy groups to test their brain activations. In the new group of 16 subjects, the high-accuracy group consisted of more female than male individuals who were experienced in landscape, whereas the low-accuracy group consisted of more males than females. In the literature review (Mohler, 2009), it was mentioned that sex and type of test affect spatial abilities, such as visualization, orientation, and rotation. Males are often better at spatial performance than females; some studies (e.g., Jansen-Osmann and Heil, 2007), however, found no sex-based differences in terms of spatial performance. That said, the present study demonstrated an opposite view, which was not the main issue affecting the results. Moreover, the tasks used in the study were selected in a pre-test performed among those experienced in landscape and novices with the intent of normalizing the difficulties in order to avoid experimental bias. Therefore, the results could reflect another view of spatial ability in landscape design, and therefore further studies are required to examine this potential issue.
Second, the results for the high-accuracy group revealed more activation in the left frontal lobe, which is responsible for high-cognition skills and a wider range of strategies in spatial ability (Mohler, 2009). In this paper, the results may be explained by considering how landscape designers think, judge, and then transform planar to cross-sections involving advanced cognitive functions in the frontal lobe, which indicates the potential function of landscape designers' brain responses using their spatial conversion capability. This statement is in line with research on spatial visualization and mental imagery in the brain related to frontal and parietal areas (Ishai et al., 2000). Moreover, from the 45 fMRI studies on creativity, it was shown that visuo-spatial creativity is induced in the right middle and inferior frontal gyrus, the left precentral gyrus, and the bilateral thalamus, which is related to the mental rotation of objects (Boccia et al., 2015). On the other hand, the result of precise positioning in a rotation mental clock found that coordinate relation is activated in the right prefrontal cortex of the right hemisphere (Trojano et al., 2002); however, this study inferred that graphic planar (2D) could be transformed into a cross-section (3D), one which is a relative position task in landscape elements. The results of the current study showed activity in the left inferior frontal gyrus of the left hemisphere, which is in line with the hemispheric lateralization in categorical and coordinate relations (Chabris and Kosslyn, 1998; Kosslyn, 1987; Kosslyn et al., 1989; Lopez et al., 2020). Furthermore, it is worth noting that the high-accuracy group included two participants who were classified as novices, which aligns with the composition of the low-accuracy group. Consequently, individuals displaying superior accuracy in spatial design, particularly those with experience, were found to engage the left inferior frontal gyrus, a region associated with proficient spatial ability. Nevertheless, it is advisable to conduct additional research to explore the neural activation patterns of both the general public and landscape designers during spatial design tasks.
Future study and suggestions
The advent of neuroscience in environmental design presents an opportunity to unveil the intricate processes of designers' brains, which have long been elusive and referred to as a "black box." Through neuroscientific approaches, researchers can now explore the neural activities involved in spatial ability thinking during design. However, the present study had limitations, including a small sample size and potential issues with data collection methods. Further research is suggested that will investigate the initial personal spatial ability in understanding the complex relationship in the context of landscape designers. Notably, a previous study by Gonen-Yaacovi et al. (2013) linked the prefrontal cortex to cognitive processes underlying creativity. Additionally, research exploring creative thinking has implicated the frontal lobe, temporal lobe, and parietal lobe as brain regions associated with creativity (Heilman et al., 2003) and their involvement in mental imagery (Pearson, 2019), which encompasses higher-level cognitive functions, including working memory, sustained attention, and flexibility, primarily governed by the frontal lobe (Dietrich, 2004).
Moving forward, prospective studies have the potential to expand our knowledge in landscape design research and neurocognition, shedding further light on the intricate workings of landscape designers' brains that lie within those above the "black box." In addition to spatial ability, landscape designers must possess crucial professional skills, such as exercising creativity in harmoniously integrating landscape elements and leveraging technological prowess. Therefore, it is imperative to discern which brain regions are activated when sketching a landscape layout on a 2D plane. Furthermore, it is essential to investigate the distinct areas of brain activation associated with manual drawing versus conceptual creativity and to explore the intricate neural mechanisms underlying the design process. These compelling inquiries could motivate researchers to further investigate the neural landscape of design thinking and to consider strategies that will enhance landscape designers' abilities by targeting the left inferior frontal gyrus.
Conclusion
The findings of this study indicate that high accuracy rates in spatial design are related to landscape expertise and associated skills, particularly those involving the engagement of the inferior frontal gyrus. This association suggests that proficient spatial ability may be a key factor influencing the design process. Conversely, the low accuracy rates observed, especially among novices, were associated with activation in the left middle occipital gyrus, indicating a reliance on visual information for spatial tasks. Furthermore, it is noteworthy that the group that achieved high accuracy rates included two novices, similar to the low-accuracy group. This implies that individuals with greater accuracy in spatial design, particularly those with experience, exhibit activation in the inferior frontal gyrus, which is associated with superior spatial ability. The findings of this study are aligned with those of previous research conducted by Tsai et al. (2021). The present study revealed that landscape designers exhibit heightened spatial ability activities in the frontal lobe. Consequently, these findings provide further evidence supporting the crucial role played by the frontal lobe in the spatial cognition of landscape designers.