Ecological Characteristics of Evergreen Forest in Yesong-ri, Wando-gun (Natural Monument No. 40)
Article information

Abstract
Background and objective
Evergreen broad-leaved forests native to the islands of the southwest coast have been adapted to a range of climatic or geographical environmental factors. Although they are of high importance as cultural and ecological landscape resources, they have been damaged by tourism and climate change, and thus measures to protect and manage them are required. This study was conducted to suggest the characteristics of the vegetation structure and management plan of the evergreen forest in Yesong-ri, Natural Monument No. 40.
Methods
Twenty-three quadrats were established in the vegetation communities where broad-leaved evergreens dominate or appear in the canopy, understory, and shrub layers, and the area where the vegetation was restored after the buildings was demolished. Based on survey data on the vegetation, the characteristics were identified after classifying the communities using TWINSPAN and DCA ordination techniques.
Results
The plant communities were classified into a total of 5 dominant groups: Castanopsis sieboldii-Camellia japonica, C. sieboldii, Celtis sinensis, Neolitsea sericea, Camellia japonica, and Machilus thunbergii-Quercus glauca. The study site was a windbreak forest that has been protected and managed by the villagers for a long time. As time passes, broad-leaved evergreens including N. sericea, Q. acuta, and C. sieboldii have flowed into and spread to the surrounding area. It seems that a similar but unique ecosystem was formed. Looking into the relationship between environmental factors and vegetation distribution, it was analyzed that the slope, clay, and Na+ had direct or indirect effects on the vegetation distribution.
Conclusion
The width of the windbreak forest was narrow, and the continuity was interrupted by the indiscriminate activities of tourists, installation of facilities, expansion and construction of warehouses and commercial facilities, and the destruction of peripheral and understory vegetation was in progress. To maintain the integrity of the forest, it is necessary to purchase and demolish buildings and facilities belonging to the interior after establishing boundaries and spheres of influence, and to restore it to a good evergreen broad-leaved forest using a reference model for areas with damaged vegetation and manage it.
Introduction
In South Korea, 9,850 ha of evergreen broad-leaved forests are estimated to remain, of which 92% are distributed in the southwest coast and islands (Kim, 2010a). Since islands surrounded by the sea can have special environmental conditions depending on their distance from the land, the distance between islands, and the area (Mologni, 2022), evergreen broad-leaved forests distributed in such places are unique vegetation adapted to the island environment (Kim et al., 2018). The geographical and environmental characteristics of islands and coasts not only enable evergreen broad-leaved forests to form different landscapes from deciduous broad-leaved forests, but also make them good models when creating Landscape and environmental forests in warm-temperate zones, increasing their utility (Oh and Cho, 1996; Park et al., 2004). However, since evergreen forests on islands and coasts are distributed in extreme areas that include steep slopes and coastal cliffs, it has been predicted that the distribution ranges will change rapidly if environmental changes such as climate change and sea level rise continue (Kim et al., 2018). Regarding the distribution of warm-temperate forests, some studies have predicted their spread due to global warming (Yoo et al., 2016), while others suggest that direct habitat destruction may slow down the spread (Park et al., 2016). Therefore, long-term monitoring of biota and vegetation changes seems to be required.
Evergreen broad-leaved communities are native to the islands of the southwest coast are recognized as valuable biological resources in terms of local landscape restoration and national-level genetic resources and biodiversity conservation strategies (Kang et al., 2022) as they have adapted to various climatic or geographical environmental factors (Park and Oh, 2002). However, as they have been destroyed by artificial influences including logging, firewood harvesting, and urban expansion, most remain in inaccessible islands, remote areas, bongsan (mountains where logging was prohibited), dangsan forests, and temple forests (Park, 2012; Kim et al., 2016; Kim et al., 2018). The damaged areas were changed into forests dominated by Quercus spp. or Pinus thunbergii, and the natural vegetation landscape of the temperate forest zone has been lost (Oh and Choi, 1993). Recently the decline in the resident population in island areas has increased the amount of Abandoned paddy, and firewood has been replaced as fuel by electricity and gas, and the damage to vegetation is decreasing (Kim et al., 2021). Despite these circumstances, since evergreen broad-leaved forests are exposed to natural and artificial hazards, it is necessary to study the process of vegetation succession as well as the change trends after external disturbances are reduced in order to set a direction for future restoration and present a reference model.
Windbreak forests on the southwest coast are also suffering a similar process of damage to that seen in the evergreen broad-leaved forests, which can be attributed to human use, poor management by administrative agencies, and lack of awareness of their importance (Park and Oh, 2002; Oh et al., 2002; Oh et al., 2007). Surveys on the vegetation structure and management status of Mulgeon-ri Windbreak Forest and Suwol Forest on the southern coast and windbreaks on the islands in Wando-gun found that excessive installation of facilities for tourism, indiscriminate use, and fragmentation caused by development are the main causes of damage to their original form (Oh et al., 2002; Oh et al., 2005; Lim et al., 2010; Lim et al., 2013; Lee et al., 2016). Windbreak forests are defined as man-made protection forests, in which trees are planted along the coast to prevent damage from tidal waves and sea winds to which the Korean Peninsula is exposed due to its locational characteristic of being surrounded by the sea on three sides (Lim et al., 2012; Oh et al., 2005). Windbreak forests are located between the sea and the land, so they not only perform a buffer or Calamity prevention function to protect houses and farmland from coastal disasters, but also play a very important role as a habitat where sea and land Life coexist (Oh and Cho, 1996; Oh et al., 2007; Lim et al., 2010). In fact, A study has reported that coastal forests made the villages in their protection range less susceptible to typhoon damage than villages without them, and greatly reduced the speed and destructive power of earthquakes and tsunamis (Lim et al., 2012).
In addition to disaster prevention and biological values, It has been part of the history of the village, so they retain traditional factors with various historical and cultural meanings (Hwang and Kim, 2015); they have high significance for tangible and intangible assets, including in village rituals held there to pray for the well-being of the village and a bountiful catch (CHA, 1993). However, despite such significance, damage to them has continued, the result of coastal reclamation, road construction, and facility installation (Lim et al., 2013; Lee et al., 2016). In particular, as windbreak forests are recognized as tourism resources, the destruction of the understory vegetation and dead trees are increasing with the establishment of trails and indiscriminate use. Since some residents also recognize windbreak forests as a factor hindering the development of their village (Lim et al., 2013), understanding their actual state in order to restore historicity and ecological soundness is an essential process for protecting windbreak forests.
Windbreak forests created in the island areas of the southwest coast are of high importance as cultural and ecological landscape resources, but are facing damage attributable to both direct factors (e.g., the activation of tourism) and indirect factors (climate change). Comprehensive measures to protect and manage them are urgently required. In Wando-gun, where the study site is located, evergreen forests in Jeongdo-ri, Yesong-ri, Maengseon-ri, and Mira-ri were designated as natural monuments due to their value as historical, cultural, and ecological resources. However, excluding those in Jeongdo-ri (Lim et al., 2010), detailed surveys of their vegetation and use have not been conducted. Therefore, this study was conducted to provide basic data on the land cover and vegetation characteristics of the Yesong-ri Evergreen Forest designated as Natural Monument No. 40.
Research Methods
Overview of the study site
Bogildo Island is located about 18 km southwest of Wando Island. With the Yesong-ri Evergreen Forest at its center, the island has peaks that include Gwangdaebong (310.5 m above sea level) to the north, Jeokjabong (425 m a.s.l.) and Mangwolbong (364 m a.s.l.) to the west. Based on the meteorological data of Wando-gun from 1991 to 2000, 2001 to 2010, and 2011 to 2020 (Korea Meteorological Administration, 1991–2020), the average annual temperature (AAT), average annual precipitation (AAP), and coldness index (CI) were 14.1–14.3°C, 1437.4–1648.3 mm, and −4.1°C·month, respectively. The distribution of Evergreen broad-leaved forest has been reported to broad-leaved forest be highly related to AAT, CI, and AAP (Yim and Kira, 1975; Yim, 1977a, 1977b; Kil and Kim, 1999; Yun et al., 2011); they are suitable for distribution in areas with an AAT of 11–15°C (Kira, 1976), an AAP of 900–1,500mm (Park et al., 2018), a CI within −4°C · month, and an average temperature of −4°C or higher in January, the coldest month (Koo et al., 2001). These criteria indicate that Bogildo Island is a suitable area for the distribution of Evergreen broad-leaved forest, which coincides with Yoo et al. (2016)’s assessment that It would be intensively distributed in coastal areas.
The Yesong-ri Evergreen Forest was built by islanders as a 1.5 km-long forest for the purpose of windbreak and disaster prevention around 220 Yesong-ri, Bogil-myeon, Wando-gun, Jeollanam-do about 300 years ago (CHA, 2019). As farmland and villages are located behind the forest, it can be assumed that it was created to protect these areas from onshore winds (Oh et al., 2005). However, after part of the southern end was destroyed by Typhoon Sarah in 1962, the Yesong-ri Evergreen Forest remains as a half-moon-shaped forest with a length of about 740 m and a width of 30 m. Windbreak forests built on the coast have not only served as a windbreak, seawall, and breakwater to protect villages and farmlands from strong sea winds and seawater but have also served as fish-shelter forests (eoburim) that provides an environment suitable for fish to live in (Kim et al., 2020). In addition, while such forests are sacred spaces to pray for the well-being of villages and a big catch of fish, they also have high ecological as well as a cultural value, as they have been well-preserved for a long time and serve as habitats for wildlife (Kim, 2010b; Lee et al., 2016). The Yesong-ri Evergreen Forest was also recognized as having these values and was designated as Natural Monument No. 40 in 1962. However, it was damaged locally and artificially, so restoration work was carried out in 2015 to bring it closer to its original shape; its historical, cultural and ecological value made it necessary to preserve and restore the coastal windbreak (Lee et al., 2020). Despite such management and restoration, it seems that it is necessary to propose conservation and management measures along with the provision of basic data, as the status of vegetation and damage were not fully understood.
Methods
To understand the ecological characteristics of the Yesong-ri Evergreen Forest, the status of use and management of the surrounding land, and the characteristics of the plant community structure of the natural monument forest were examined. The survey range of the land use status was set to a radius of 1km or more from the boundary including the forest area. By using land cover maps with mid-level categories provided by the Environmental Geographic Information Service (https://egis.me.go.kr), the vegetation type of the forest, its surrounding cultivated land, and the land cover type of the urban area were classified. The land use type was classified into mid-level categories, including a residential area, industrial area, public facilities area, transportation area, urban support facility area, bare land and construction area, cropland, wetland and river, and artificial green area. This was sub-classified into a total of 19 types of land use based on detailed characteristics. In terms of damage and management status, the location of protection and public use facilities, and buildings that break continuity in the area designated as a natural monument and surrounding areas that may affect the forest were identified.
To determine characteristics including species composition and stratification of vegetation constituting the forest, a total of 23 quadrats (aka square plots) were determined, including 22 areas dominated by evergreen broad-leavedor deciduous broad-leaved trees, and one restored area (Fig. 1). The first reason why a survey plot was set in the restored area where evergreen broad-leaved had been planted after demolishing buildings was the symbolism of restoring a natural monument. The second reason was to present a management direction for the site to be restored after the damage or deterioration factors are removed from the long-term perspective. Therefore, despite its narrowness, a quadrat was set up to understand the planted species and stratification. Each plot was based on 10 m × 10 m (100 m2), and 1–4 plots were installed together in consideration tree species and height, and degree of damage. Based on the vegetation survey data, the plant communities were classified using two-way indicator species analysis (TWINSPAN), a method for the hierarchical divisive classification of communities, and detrended correspondence analysis (DCA), an ordination method, and then each characteristic was determined (Hill, 1979). The target site was entirely surveyed since a forest was created to the south, and a natural forest dominated by Castanopsis sieboldii remained on the steep slope to the north. In setting the survey plots, environmental factors including slope, Aspect, and elevation above sea level, along with whether the canopy layer, understory layer and shrub layer were formed, were considered. Targeting points with similar dominant species in the canopy layer, quadrats (square plots) were set in consideration of the distribution of and damage to understory vegetation, and then the quadrat method was applied to identify plant species names for each stratum and measure the size. In general, evergreen broad-leaved forests distributed in imperfectly or perfectly preserved areas mostly form three strata of vegetation (Lalzarzovi and Tluanga, 2017). The classification was made by considering this. The tree species in each quadrat were classified based on the location of the crown into the canopy layer receiving direct sunlight, the intermediate understory layer, and the shrub layer with a height of 2 m or less, and the species name and size were identified and recorded. As for the size, the diameter at breast height (DBH) was recorded for trees and understory layers with a DBH of 2 cm or more, and the crown width for the shrub layer with a tree height of 2m or less.
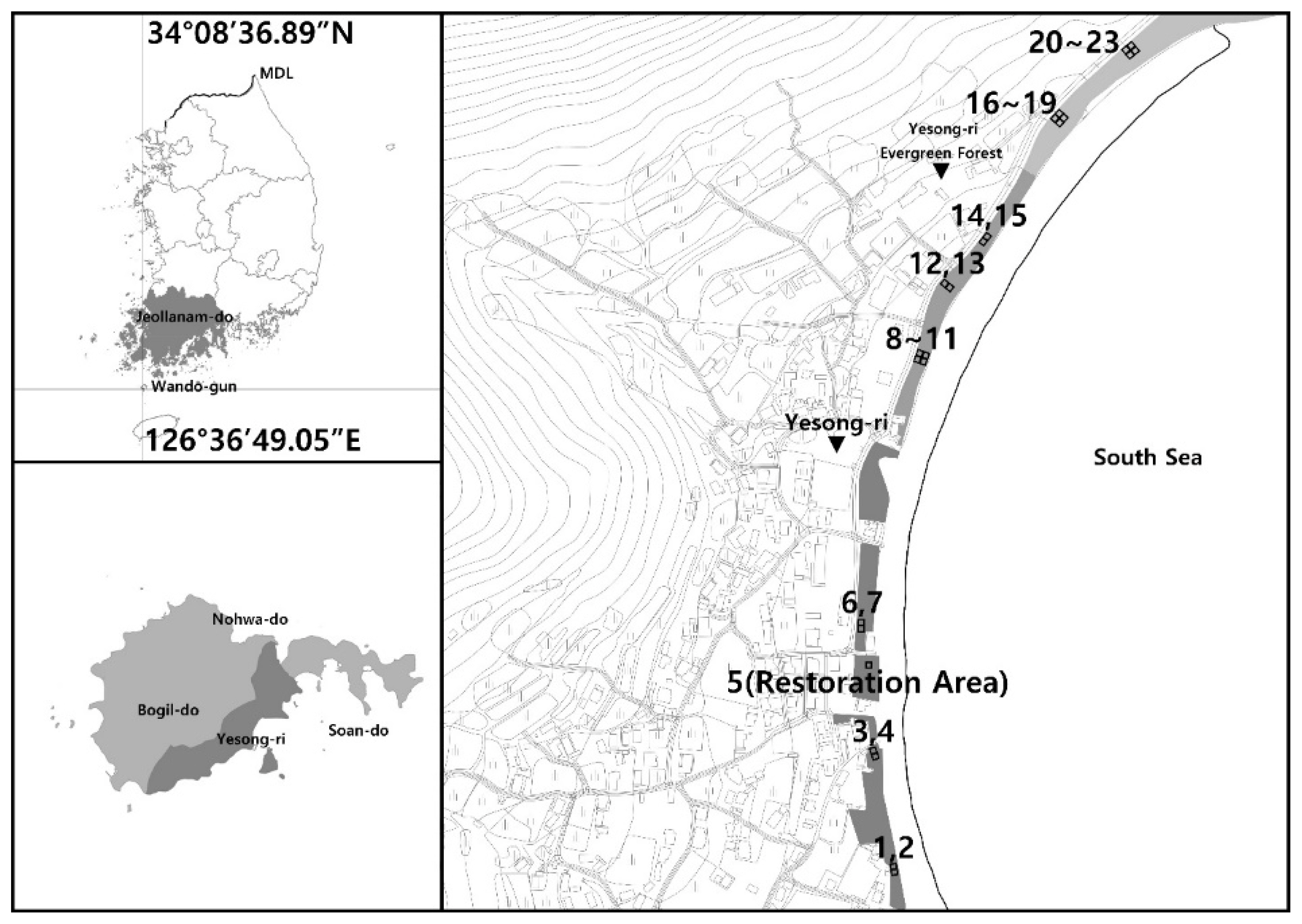
Location map of the study site and plant community survey plots at Yesong-ri.
To compare the relative dominance of tree species in the survey plots, by integrating the importance percentage (IP) of Curtis and Mclntosh (1951), IP, the relative dominance expressed as a percentage, was analyzed by the crown stratum (Brower et al., 1977). IP was calculated as (relative density + relative coverage)/2 for each species. The mean importance percentage (MIP) was calculated as {(canopy layer IP×3)+(understory layer IP×2)+(shrub layer IP×1)}/6 weighted by crown stratum considering the relative height of each individual tree (Park, 1985). For measures representing the various degrees of species composition by community, species diversity (H′), evenness ( J′), and dominance (D′) were calculated by applying Shannon’s formula (Pielou, 1975), and similarity index (SI) was calculated using the formula of Whittaker (1956). For the number of species and population by the community, the average value was estimated for communities where two or more survey plots were set based on a plot area of 100m2. To determine the physicochemical characteristics of the soil, samples were taken from the B layer after removing the organic matter layer, and soil pH, organic matter content, cation-exchange capacity, effective phosphate, and electrical conductivity were analyzed (National Academy of Agricultural Science, 2000). In addition, a detrended canonical correspondence analysis (DCCA) was conducted to identify the relationship between vegetation and environmental factors; like DCA, the distortion of the second axis can be improved using canonical correspondence analysis (CCA; Lee et al., 2014), so DCCA, one of the ordination methods, was used in the Multivariate Statistical Package (MVSP) version 3.2. As for management, the application of evergreen broad-leaved forest restoration models and long- and short-term planning directions for the windbreak forest itself and the affected areas were presented.
Results
Land use
According to the land use survey results(Fig. 2, Table 1), the area designated as a natural monument was 58,486m2, but the forest area was 14,753m2, about one-third of that area, as the natural monument includes the gravel coast and sea surface. The surrounding area consisted of natural greenery (40.73%) and sea surface (41.05%), and was connected to the sea to the east of the forest area and to the forest through villages to the west. Villages, which have been formed to the west of the forest, consisted of residential areas (detached houses), commercial areas including rental cottages, and farmland. Most of the areas that had been used for agricultural fields in the past were now in the form of fallow land where agricultural activities had been abandoned, or bare land where fishing gear was loaded.
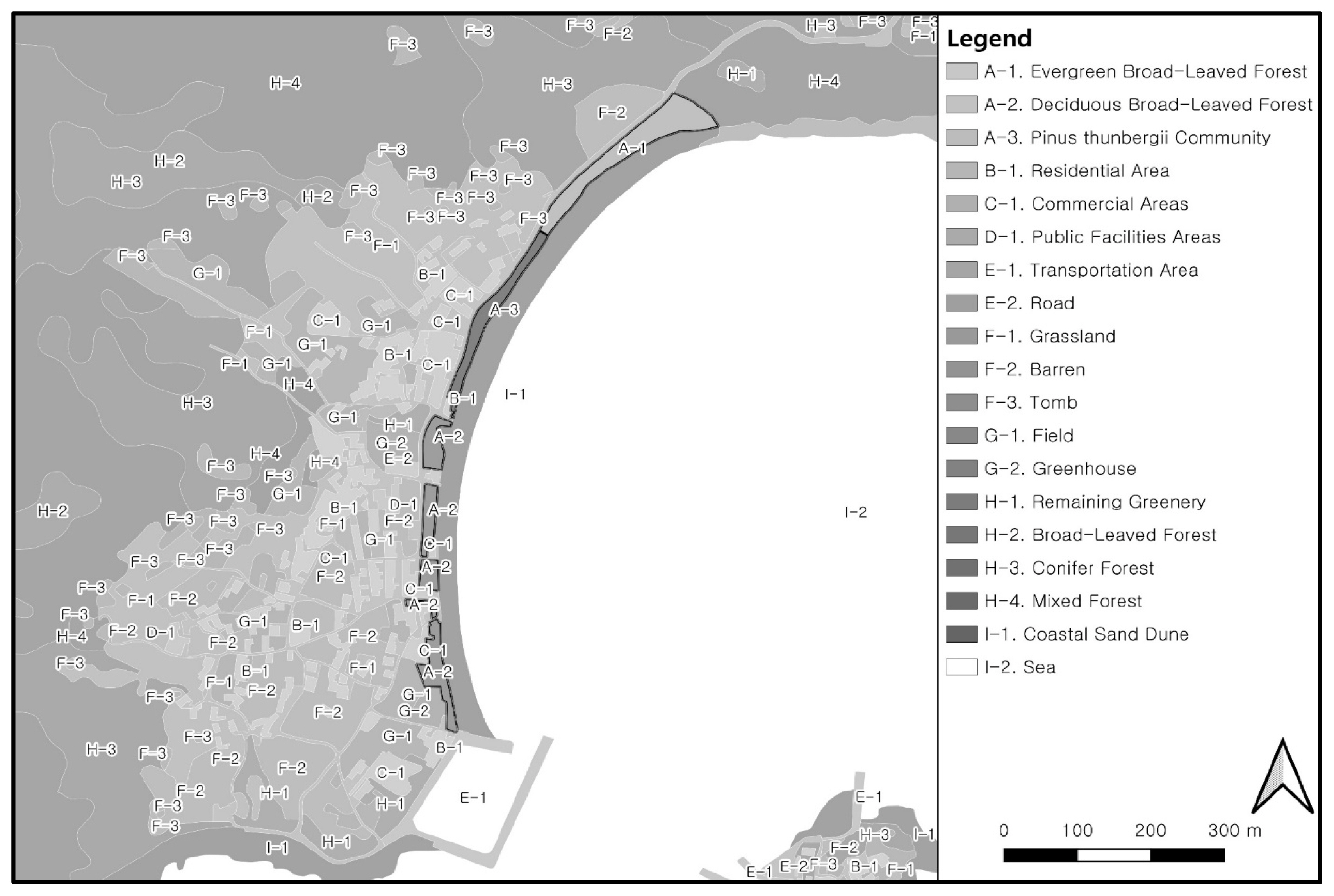
Land use pattern around Yesong-ri evergreen forest.
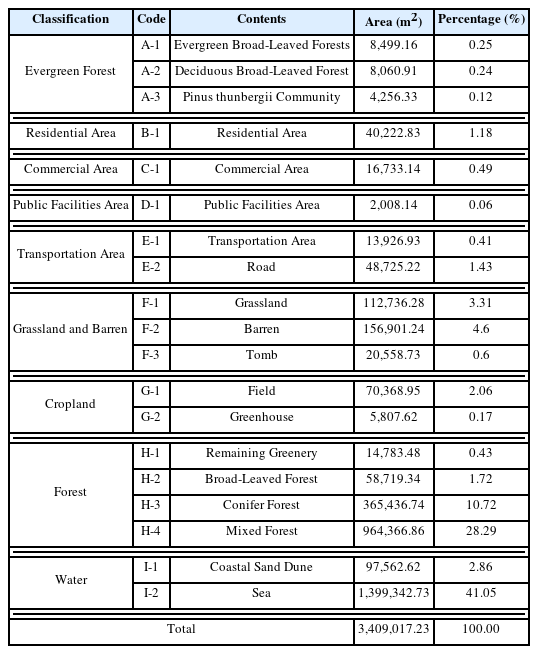
Area and percentage by type of land use pattern around Yesong-ri evergreen forest
Ecological characteristics of vegetation
Community classification and general overview
Plant communities of Yesong-ri Evergreen Forest, a natural monument, were classified using DCA and TWINSPAN techniques. In the dendrogram of DCA classification, the eigenvalues of the first and second axes were 0.55621 and 0.32925, respectively, showing about 90% explanatory power, which was diagrammed based on the two axes (Fig. 3). A total of 5 dominant groups were classified: evergreen broad-leaved forest (natural forest) and Castanopsis sieboldii on the left side of the upper part, and a group in which Pinus thunbergii and broad-leaved evergreens compete on the right; and Pinus thunbergii on the left of the lower part, and Celtis sinensis on the right. The area was small, but the vegetation restored by planting Machilus thunbergii and Quercus glauca was classified separately. For the dendrogram of TWINSPAN ordination (Fig. 4), in Division I, groups in which Machilus thunbergii appeared and those in which it did not were divided. The latter has been restored by planting Machilus thunbergii and Quercus glauca, etc. after demolishing the buildings. In Division II, the groups in which Castanopsis sieboldii appeared were classified into Castanopsis sieboldii communities where Quercus acuta was an indicator species and Castanopsis sieboldii-Camellia japonica communities where Quercus acuta was not. In Division III, the groups in which Castanopsis sieboldii did not appear were classified into Pinus thunbergii communities where Elaeagnus macrophylla was an indicator species, Neolitsea sericea communities where Camellia japonica was an indicator species, and Celtis sinensis communities where Camellia japonica was not.

Dendrogram of 23 sites classified by DCA classification. ●: Castanopsis sieboldii, ▴: Pinus thunbergii, ▪: Celtis sinensis, ◆ Pinus thunbergii, Evergreen Broad-leaved, ◈: Machilus thunbergii-Quercus glauca
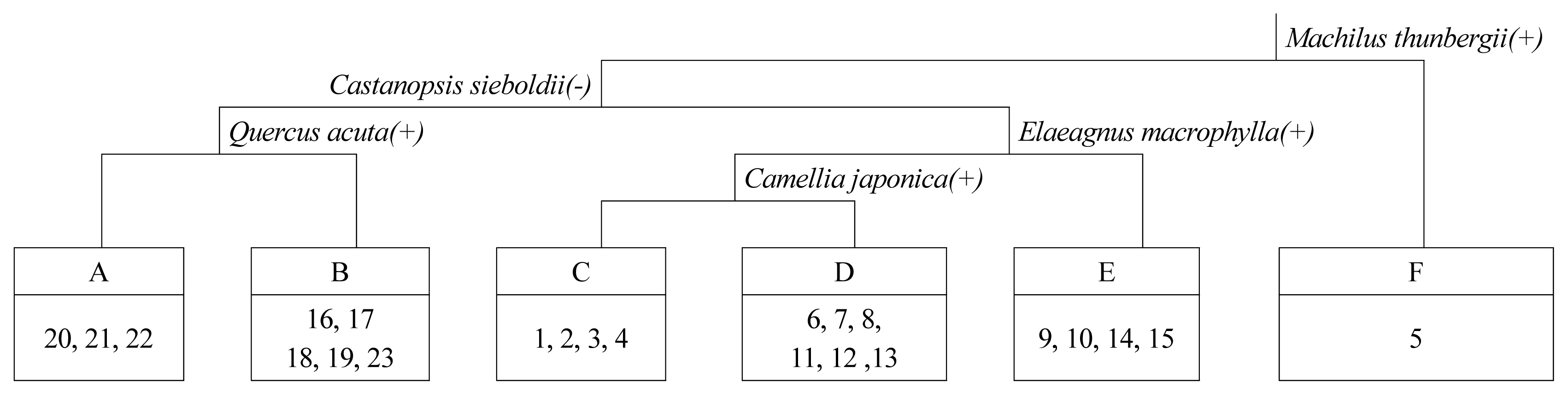
Dendrogram of 23 sites classified by TWINSPAN ordination. A: Castanopsis sieboldii-Camellia japonica, B: Castanopsis sieboldii, C. Celtis sinensis, D: Neolitsea sericea, E: Pinus thunbergii, F: Machilus thunbergii-Quercus glauca.
For community classification using DCA and TWINSPAN techniques, forests dominated by Castanopsis sieboldii on steep slopes were similar except for the aspect that Castanopsis sieboldii communities and Castanopsis sieboldii-Camellia japonica communities were classified depending on whether Quercus acuta was an indicator species. The Yesong-ri Evergreen Forest was classified into Castanopsis sieboldii community, Castanopsis sieboldii-Camellia japonica community, Neolitsea sericea community, Pinus thunbergii community, Celtis sinensis community, and damaged Machilus thunbergii-Quercus glauca community depending on whether Evergreen broad-leaved trees of the canopy layer dominate and by indicator species.
Table 2 shows the topographical characteristics of six communities classified by DCA and TWINSPAN, and the status of vegetation by stratum. The Castanopsis sieboldii-Camellia japonica community was connected to the windbreak forest, but it was a remaining natural forest that avoided damage as it was located on a steep slope of 38°. The Castanopsis sieboldii community had similar location conditions to the Castanopsis sieboldii-Camellia japonica community, but was distinguished from the latter by the indicator species and located in the southwest direction with 22.4m above sea level and a slope of of 28.3°. The Celtis sinensis community, which is vegetation distributed in the flatland facing northwest at 19m above sea level and a slope of 9°, was good in the understory layer (coverage 60%) and shrub layer (coverage of 40%), but in the canopy layer was in a state of withering due to damage from the sea wind (coverage of 35%). The Neolitsea sericea community was distributed in the southwest at 20.7m above sea level and a slope of 8.7°, and its coverage was good on all strata. The Pinus thunbergii community was located adjacent to a place where broad-leaved evergreens were distributed, but the coverage by stratum was good. In 2015, Machilus thunbergii and Quercus glauca were planted on the flatland where the buildings were demolished.
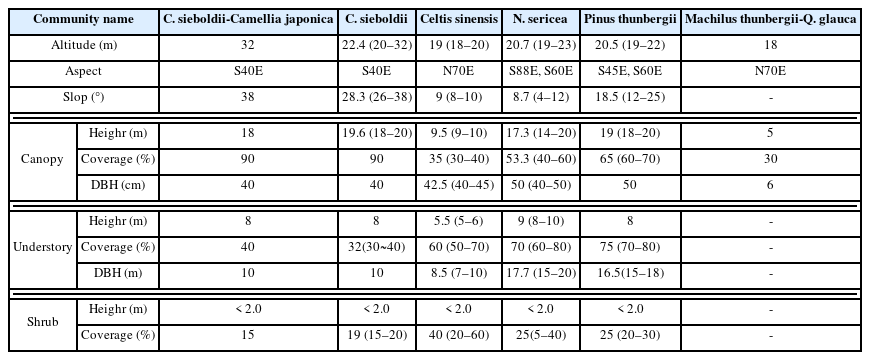
General description of the physical status and vegetation of the surveyed site
Importance percentage and species status
Table 3 shows the relative dominance (IP) and average relative dominance (MIP) of the major species within the communities. For the Castanopsis sieboldii-Camellia japonica community, Castanopsis sieboldii (IP: 85.1%) was the main species in the canopy layer, Camellia japonica (IP: 77.6%) in the understory layer, Mallotus japonicus (IP: 39.4%) and Kadsura japonica (IP: 21.9%) in the shrub layer. For the Castanopsis sieboldii community, Castanopsis sieboldii (IP: 85.9%) and Quercus acuta (IP: 11.6%) competed in the canopy layer, and Camellia japonica (IP: 32.7%) and Castanopsis sieboldii (IP: 24.2%) in the understory layer, Hedera rhombea (IP: 18.7%) and Rubus hirsutus (IP: 13.9%) in the shrub layer. For the Celtis sinensis community, Celtis sinensis (IP: 91.5%) was dominant in the canopy layer, Litsea japonica (IP: 62.9%) and Neolitsea sericea (IP: 16.9%) in the understory layer, and Hedera rhombea ( IP: 38.2%) in the shrub layer. In the canopy layer, Celtis sinensis, a deciduous broad-leaved tree, was dominant, but its coverage was low at 30–40%, and seems to have been damaged by use or Sea breeze. For the Neolitsea sericea community, Neolitsea sericea (IP: 36.8%) was the dominant species in the canopy layer; Camellia japonica (IP: 41.9%), Litsea japonica (IP: 28.2%), and Neolitsea sericea (IP: 21.3%) in the understory layer; and Neolitsea sericea (IP: 32.4%) and Hedera rhombea (IP: 22.2%) in the shrub layer. For the Pinus thunbergii community, Pinus thunbergii (IP: 70.7%) and Pinus densiflora (IP: 17.7%) appeared in the canopy layer, Litsea japonica (IP: 23.8%), Neolitsea sericea (IP: 23.4%), and Ligustrum japonicum (IP: 21.7%) in the understory layer, and Hedera rhombea (IP: 15.1%), Sasa borealis (IP: 14.6%), and Litsea japonica (IP: 12.5%) in the shrub layer. In the Machilus thunbergii-Quercus glauca community that was restored after demolishing the existing buildings, only Machilus thunbergii (IP: 63.8%) and Quercus glauca (IP: 36.2%) were planted.

Importance percentage of major woody plants by stratum in 6 communities
Looking into the number of species and population by the community (Table 4), the number of species was 4–20 for each survey plot, excluding the restored Machilus thunbergii-Quercus glauca community (2 species). The average number was 7–13.8 species for each community, and the Castanopsis sieboldii community, Celtis sinensis community, and Pinus thunbergii community had 10 species or more. It seems that the reason why the number of species for each survey plot did not show significant differences was that the Yesong-ri Windbreak forest had been exposed to similar environmental conditions and disturbances. The population per 100 m2 was 2.5–5.8 in the canopy layer, 9.4–21.8 in the understory layer, and 54.7–157.3 in the shrub layer, indicating differences according to the intensity of use. The population of the Pinus thunbergii community was the largest with 5.8 in the canopy layer, followed by the Castanopsis sieboldii-Camellia japonica community and the Castanopsis sieboldii community with 4 or more each, and the other communities with 2.5. In the understory layer, 18 or more were observed for Neolitsea sericea community, Pinus thunbergii community, and Celtis sinensis community, but less than 18 for the remaining communities. In the shrub layer, the Neolitsea sericea community had the highest population with 157.3, and the Castanopsis sieboldii-Camellia japonica community had the lowest population with 54.7, while the remaining communities had 96–125. The Machilus thunbergii-Quercus glauca community, which was damaged, had only 22 trees planted in total, which accounted for 30% of the population in the Castanopsis sieboldii-Camellia japonica community, which had the smallest population. Not only the number of species in the damaged area but also the population were found to be small.

Mean number of species and population of six communities
Diversity and similarity
Species diversity for each community ranged from 0.2847 to 0.9237; the Castanopsis sieboldii community (0.9237), Pinus thunbergii community (0.8110), and Celtis sinensis community (0.7743) had high species diversity, while in the Castanopsis sieboldii-Camellia japonica community (0.6456) and Neolitsea sericea community (0.5973) species diversity was relatively low. The Machilus thunbergii-Quercus glauca community (0.2847), which was damaged, showed the lowest species diversity, with only two species planted (Table 5).
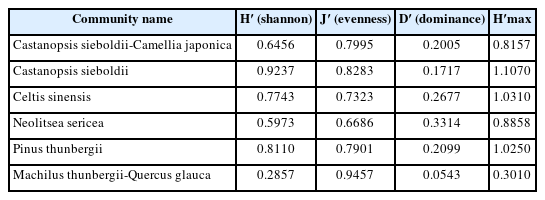
Species diversity indices of six communities
In terms of similarity between communities, the similarity between the Castanopsis sieboldii-Camellia japonica community and Castanopsis sieboldii community was high at more than 60%. This is because the native vegetation dominated by Castanopsis sieboldii remained in the slope area and was well preserved. In addition, the Neolitsea sericea community and Pinus thunbergii community showed a similarity of more than 45%, which seems to be because the location conditions including elevation and slope were similar, and the succession of the warm-temperate forest zone was taking place to the middle stage. On the other hand, the similarity index between the Machilus thunbergii-Quercus glauca community created in the damaged forest area and other communities were heterogeneous, at less than 10% (Table 6).

Similarity index among 6 communities
Soil characteristics
By examining soil characteristics (Table 7), it was found that soil acidity (pH) ranged from neutral to subacid with an average of 5.59–6.43 in all the communities, excluding alkaline (pH 7.91) in the damaged area. Cation exchange capacity was 0.7–1.6cmol/kg for K+, 9.3–39.0 cmol/kg for Ca9.3-, 1.1–10.8 cmol/kg for Mg2+, and 0.2–1.3 cmol/kg for Na9.3-, which was the lowest in the damaged area. Ca9.3- was relatively low in the Castanopsis sieboldii-Camellia japonica and Castanopsis sieboldii communities remaining on steep slopes. The electrical conductivity ranged from 0.197 to 0.686 dS/m on average, and the Neolitsea sericea community (0.631 dS/m) and Celtis sinensis community (0.625 dS/m) showed high electrical conductivity. The organic matter (OM) content ranged from 0.83 to 3.09% on average; excluding the damaged area, the Castanopsis sieboldii community had the lowest OM at 1.87%, and the Pinus thunbergii and Neolitsea sericea communities, which were known to adapt well to the sea wind, had relatively high OM at 3.09% and 2.96%, respectively. The Castanopsis sieboldii-Camellia japonica and Castanopsis sieboldii communities, which had an average slope of more than 20°, were found to have an available phosphate of around 200 mg/kg. The Pinus thunbergii community (298.9 mg/kg), Neolitsea sericea community (482.8 mg/kg), and Celtis sinensis community (422.9 mg/kg) seemed to be under management because of their high available phosphate content.
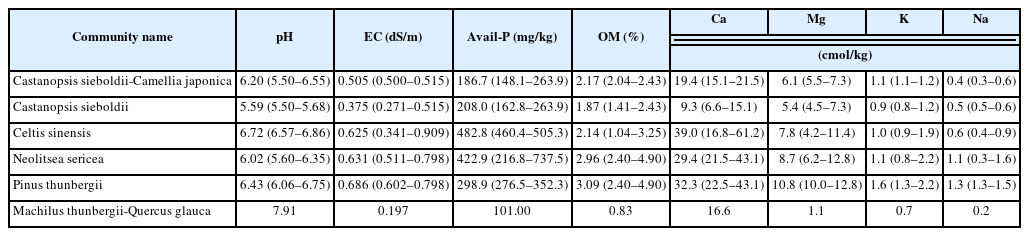
Soil physics and chemistry characteristics of 6 communities
Relationship between environmental factors and vegetation
The results of DCCA ordination with environmental factors and 22 survey plots excluding the damaged area were diagrammed based on the first and second axes, where the eigenvalues are concentrated at 80% or more (Fig. 5). Of 13 environmental factors, 12, including soil pH, OM content, electrical conductivity, elevation, slope, soil physics and cation exchange, were found to have an effect on vegetation distribution. The Castanopsis sieboldii community was located on steep slopes, with high clay and silt content. The Pinus thunbergii community was distributed in an area with high OM content along with high elevation, while the Celtis sinensis community was distributed in an area with high pH. Castanopsis sieboldii-Camellia japonica and Neolitsea sericea communities appeared in areas with high available phosphate, Ca2+, electrical conductivity, Mg2+ and Na+. In summary, it was analyzed that slope, elevation, pH, and clay among physical properties, and Na+ directly or indirectly affect vegetation distribution.
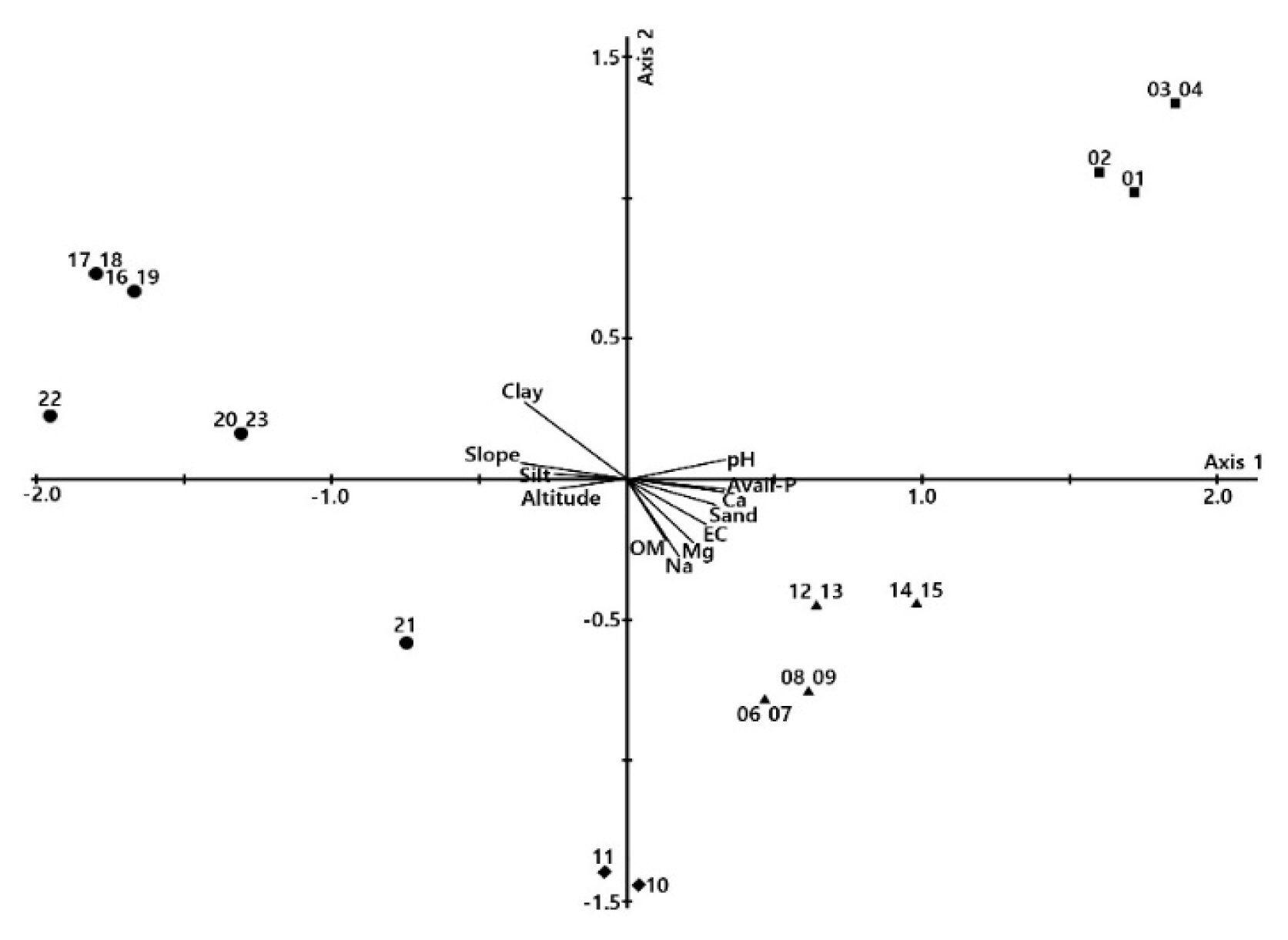
A DCCA ordination diagram of vegetation communities respect to environmental variables at Yesong-ri. ●: Castanopsis sieboldii, ▴: Camellia japonica-Neolitsea sericea, ▪: Celtis sinensis, □: Pinus thunbergii.
Status of deterioration and management history
Management of Yesong-ri Evergreen Forest, a natural monument, seems to have begun with the construction of an embankment to protect it in 1966 as part of recovery efforts following the damage caused by Typhoon Sarah in 1963 (National Research Institute of Cultural Heritage, 2011). Afterwards, maintenance work was carried out, including repairing the embankment in 1978 and 1984, constructing a new public toilet in 1986, and installing a fence in 1997. Regarding the damage to the forest, it seemed that no resolute protection measures were taken given that new houses were built in its environs in 2005 and Fish farms were expanded in 2010; in addition, rental cottages and warehouses were continuously expanded or built between the windbreak forest and the villages. In 2012, fires from construction wastes loaded near the forest spread to the protected area, causing great damage, but it seems that no practical management was performed (Korea Forest Research Institute, 2015). After demolishing the buildings built inside the forest in 2015, the damaged area was restored by planting Machilus thunbergii and Quercus glauca.
Facilities installed inside the forest included a notice announcing that it was designated as a natural monument and other information boards, fences for protection, trails that traverse the inside, public toilets, and drinking fountains (Fig. 6). In particular, the public toilets and shops built inside the forest area were considered to have an effect on fragmentation in terms of the movement of wildlife as well as the landscape.
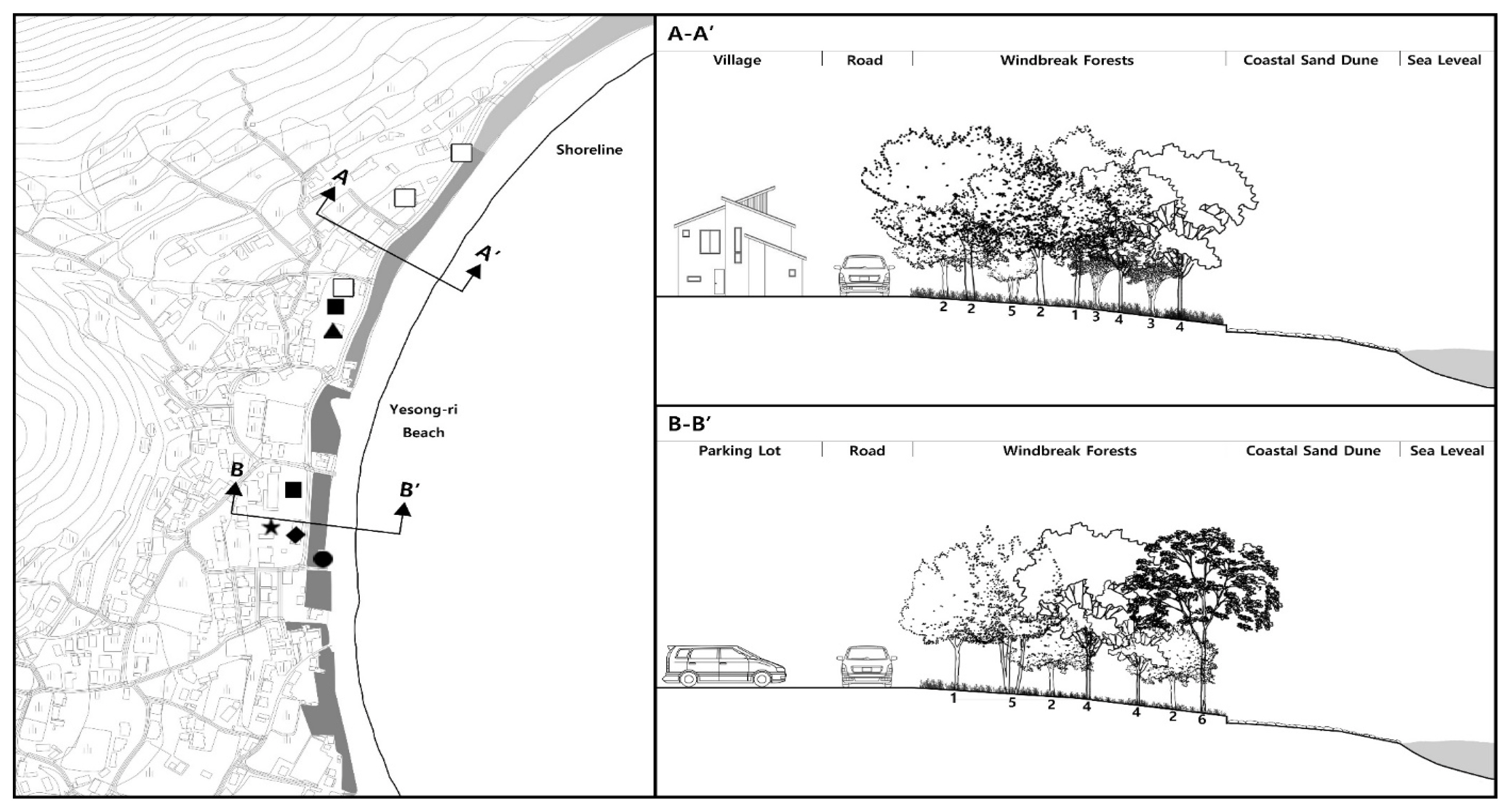
Cross Section of A-A′ and B-B′ at Yesong-ri Evergreen Forest. ●: Public Toilet, ▴: Public Shower Room, ▪: Parking Lot, □: Sewage Treatment Plant, ◈: Commercial Area, ★: Fishery Material Storage Area. 1: Machilus thunbergii, 2: Neolitsea sericea, 3: Litsea japonica, 4: Pinus thunbergii, 5: Camellia japonica, 6: Pinus densiflora.
Despite the installation of wooden fences along the boundary and walking decks inside the forest, acts including not using the designated trails or setting up tents inside the windbreak forest during peak season were spreading the damage to the understory vegetation. Since the forest is located within a national park, basic amenities such as toilets and drinking fountains are taken for granted, but these were breaking the continuity along with houses and shops. Looking at the cross-section (Fig. 6), the north area (A-A′) was indirectly connected to residential or commercial facilities, roads, and windbreak forest, so there seemed to be no damage due to building extensions and renovation. On the other hand, the south area (B-B′) was directly connected with parking lots, commercial facilities, residential areas, warehouses without a buffer space, which not only formed poor scenery, but also damaged the vegetation of the natural monument due to building expansion and renovation.
The community in which Castanopsis sieboldii is the dominant species located on the steep slope seemed to be undamaged, but the vegetation on the flatland adjacent to the beach was damaged by various natural and artificial factors, including sea wind and land use. Fig. 7 showed representative damaged communities. Site-02 (Left) is a Celtis sinensis community, where although walking decks were installed at the lower part, the understory vegetation was being destroyed by human trampling caused by indiscriminate access for camping, relaxation, and walking during the summer peak season. Site-05 (Right) was restored after the existing buildings were demolished, but other than Machilus thunbergii and Quercus glauca, understory and shrub vegetation were not introduced, so it was in a different state from the other communities.

Floor plan in Site-02 (left: Celtis sinensis) where the understory vegetation was damaged and Site-05 (right: Machilus thunbergii-Q. glauca) where only canopy trees were planted at Yesong-ri Evergreen Forest. 1: Celtis sinensis, 2: Litsea japonica, 3: Neolitsea sericea, 4: Quercus glauca, 5: Machilus thunbergii, 6: Hedera rhombea, 7: Pittosporum tobira, 8: Elaeagnus glabra.
Regarding the planting density for future restoration, only species of canopy vegetation with 22 individuals of 2 species were planted in the damaged area (Machilus thunbergii-Quercus glauca community), whereas it was analyzed that vegetation in the windbreak forest had 7–13.8 species with 2.5–5.8 individuals in the canopy layer, 9.5–21.8 in the understory layer, and 54.7–157.3 in the shrub layer. It would be desirable to use this windbreak as a model to supplement the planting species and population of the damaged area. Even in the soil environment of the damaged area, which had a pH of 7.91 and an organic matter content of 0.83%, it was considered that improvement was needed since the base for plant growth was poor compared to other communities in the windbreak forest as well as the forest.
Discussion
It has been suggested that village forests created on the southwest coast are differentiated from forests created inland as cultural landscapes that recognize the abundance and value of coastal villages and represent their identity (Lim et al., 2012). Despite the high value of coastal windbreak forests in terms of ecology, culture, and disaster prevention, the installation of amenities to promote tourism and damage caused by development around them is a common phenomenon (Lim et al., 2013; Lee et al., 2016). In addition to Evergreen broad-leaved trees, Pinus thunbergii and Celtis sinensis are growing at the Yesong-ri Evergreen Forest, a natural monument. Residents living behind the forest designated a large tree as a guardian tree, and since then have held a ritual for sea gods on April 12 of the lunar calendar, which has the nature of a village ritual to pray for the well-being of the village and a bountiful catch; they are thus striving to protect them as important tangible and intangible assets of the village (CHA, 1993). Despite such efforts by the residents, the need for protection was increasing as damage to the understory vegetation was being aggravated due to digging and collecting Machilus thunbergii and Camellia japonica out of the ground, and camping by tourists (Kim and Oh, 1997). Since 2015, the damaged area has been restored close to its original form, but the results have been insignificant. Therefore, this study was conducted, as it is necessary to accurately diagnose the vegetation and damage in order to protect and manage the Yesong-ri Evergreen Forest.
To understand the ecological characteristics of the windbreak forest, the status of land use within its sphere of influence and the plant community structure of major vegetation were examined. Based on a land use survey, in the north, the forest area was separated from the urban area by a road, but in the south, it was directly connected with the urban area and damaged by the extension and reconstruction of buildings. Although the forest is located within a national park, it suffered damages that included fragmentation of the forest area by the amenities of the beach, such as public toilets and shops, or the cutting of the crowns of old big trees. The vegetation was classified as follows: Castanopsis sieboldii, Castanopsis sieboldii-Camellia japonica, Neolitsea sericea communities, which are Evergreen broad-leaved trees widely distributed in islands and coastal areas of Dadohaehaesang National Park (Kim et al., 2021); Pinus thunbergii community, which mainly appears in coastal forests and windbreak forests (Park et al., 2009; Lee et al., 2010); Celtis sinensis community widely distributed along the coast due to its high tolerance to salt spray drift (Kim, 2010a); and restored Machilus thunbergii-Quercus glauca community. The distribution of vegetation was found to be directly or indirectly affected by environmental factors including slope, elevation, pH, and clay among the physical properties, and Na+. Studies related to Bogildo Island reported that vspecies including Cinnamomum yabunikkei, Neolitsea sericea, Camellia japonica, Quercus acuta, and Castanopsis sieboldii as well as Pinus thunbergii and Celtis sinensis coexisted or competed (Kim and Oh, 1997; Kim et al. 2021), which were also similar to the analysis results in this study. Consequentially, the evergreen broad-leaved forest constituting the Yesong-ri windbreak forests seems to have formed a similar but unique ecosystem to the surroundings on the results of the natural inflow from the surroundings and the succession of the warm-temperate forest zone.
Looking into the characteristics of each community, the Castanopsis sieboldii community distributed in coastal steep slope areas at 15–91 m above sea level (Lee, 2022; Kang et al., 2016) showed similar characteristics to the counterpart communities on the southwest coast, where Castanopsis sieboldii had a high coverage in the canopy layer, and broad-leaved evergreens including Camellia japonica, Machilus thunbergii, and Cinnamomum japonicum Siebold dominated in the understory layer (Han et al., 2007; Cho et al., 2017; Lee et al., 2020; Kim et al., 2021; Lee, 2022). As most studies have reported Castanopsis sieboldii as a species emerging at the climax of the succession (Park et al., 2018; Lee et al., 2019; Lee, 2022), this status would persist if there’s no disturbance. In the Neolitsea sericea community, where various Evergreen broad-leaved species characteristically grow in a jumble in the lower layer (Kim et al., 2005; Lee et al., 2019), Neolitsea sericea tended to become dominant toward the lower layer. According to Yun et al. (2014), remaining habitats of Neolitsea sericea are highly likely to spread or move north with climate change, so the current status, which is the middle stage of the succession series, will be maintained. In the Pinus thunbergii community, Neolitsea sericea, which belongs to the middle stage of the succession series of warm-temperate forests, appeared in the lower layer (Park et al., 2018), but the current status will be maintained as Pinus thunbergii dominates in the canopy layer. Since the succession of evergreen conifers in the warm-temperate forest zone progresses through Neolitsea sericea and Machilus thunbergii (middle stage) to Castanopsis sieboldii and Quercus acuta (Park et al., 2018; Lee et al., 2019; Lee, 2022), the community will change into one dominated by Neolitsea sericea with high coverage in the understory vegetation.
Most windbreak forests on the southwest coast have a structure in which deciduous broad-leaved trees including Zelkova serrata, Celtis sinensis, and Chionanthus retusus are dominant rather than Evergreen broad-leaved trees (Lim et al., 2010; Lim et al., 2013; Lee et al., 2016). Although deciduous windbreak forests in the coastal areas have high protection value in terms of ecology and local cultural history (Oh and Kim, 1997), they have suffered serious damage from external factors including sea wind, use, and human trampling (Lee et al., 2016). The Celtis sinensis community, located between the bathing beach and the urban area, was also in need of active protection as trees in the canopy layer had low coverage and were dying due to the influence of human trampling and sea Breeze. However, since the coverage of the understory and shrub layers is high (50–70%) and Neolitsea sericea and Machilus thunbergii are appearing, if the community is protected and managed, the vegetation will be restored to an evergreen broad-leaved forest. In the Machilus thunbergii-Quercus glauca community created in the area restored after demolishing the existing buildings, only Machilus thunbergii (IP: 63.8%) and Quercus glauca (IP: 36.2%) have been planted. Since Bogildo Island belongs to the warm-temperate forest zone, it is desirable to plant Machilus thunbergii and Quercus glauca, which are in the middle stage of the succession process, but the restoration of the community seems to be delayed as the understory vegetation has not been added.
Considering the findings that evergreen conifers and deciduous broad-leaved trees among vegetation distributed in a warm-temperate forest zone are changed to Evergreen broad-leaved trees in the succession series (Park et al., 2018; Lee et al., 2019), the community dominated by Pinus thunbergii, and the Celtis sinensis community disturbed by external factors, will be changed into communities dominated by Neolitsea sericea and Machilus thunbergii appearing in the understory vegetation. However, as the community in which species in the middle or climax stage of succession, such as Neolitsea sericea and Castanopsis sieboldii, appear in the canopy and understory layers, is maintaining the status quo (Park et al., 2018; Lee, 2022), evergreen broad-leaved forests will be maintained if artificial disturbance factors are managed. On the other hand, if global warming progresses, the area of evergreen broad-leaved forest can increase, or move north (Yun et al., 2011; Yun et al., 2014; Yu et al., 2020), and Pinus thunbergii and deciduous broad-leaved forests that include Celtis sinensis are also highly likely to change to evergreen broad-leaved forests, so climatic factors should be considered in selecting restoration communities. Since soil properties may be changed depending on the dominant vegetation, and the soil itself has a great effect on the distribution, growth and renewal of forest trees (Park et al., 1990), there was a large difference between the remaining forest and the restored area.
To prevent damage to and restore the coastal windbreak forests, which are an ecological, historical, and cultural heritage that has a shared history with coastal villages, it is necessary to combine legal and institutional measures with direct management (Oh et al., 2007; Lim et al., 2013). To fully manage the Yesong-ri Evergreen Forest, it is necessary to set up areas that physically affect the forest, and in the short term, install fences around it and then prohibit access, close internal trails and walking decks, and remove benches and sports facilities. From a long-term perspective, it is necessary to suggest ways to detour roads that interrupt the continuity of the forest, purchase and demolish certain buildings and facilities, and minimize the impact of building extensions and renovations.
Direct management should be involved in supplementing the damaged vegetation in the forest, nurturing next-generation trees for restoration after the demolition of facilities, and restoring the understory vegetation (Park et al., 2004; Lee et al., 2016). Since it is important to form a multi-layered structure of vegetation to ecologically and functionally stabilize windbreak forest (Kim et al., 2012), prohibiting camping and relaxation in the forest and establishing a multi-layered structure should be priorities for vegetation management and restoration. Park et al. (2018) and Lee et al. (2019) reported that the vegetation of islands distributed in the warm-temperate forest zone changes to evergreen broad-leaved trees in the succession series; it would be desirable to select a restoration model in consideration of the species and density of evergreen broad-leaved communities in the remaining forest. On the other hand, species diversity was reported to be high when various plant species adapt to the microgeography and microclimate over a long period of time and form an ecologically stable vegetation structure (Kim, 2010a), but in this study, with the exception of Castanopsis sieboldii, it was lower than Mulgeon-ri Windbreak Forest (0.9619; Lee et al., 2016) and Suwol Forest, Tongyeong (0.8273; Lim et al., 2013). This seems to be because the width of the windbreak forest was narrow and the bathing beach was located there, making access easier. In the study site, as damage to vegetation including the destruction of understory vegetation caused by indiscriminate access by tourists has increased (Kim and Oh, 1997), it seems that good vegetation will be maintained or restored only when continuous management and restoration are performed considering the succession series. To restore coastal windbreak forests, it is important to select plants with high tolerance to soil salinity and salt spray drift as well as the distribution potential of vegetation (Kim, 2010a). Therefore, the planting of Quercus glauca and Machilus thunbergii in the area where the buildings were demolished seems appropriate, considering that a literature review or a survey of surrounding forests found that broad-leaved evergreens had appeared there. However, since a multi-layered structure of vegetation is suitable for windbreak forests to achieve ecological and functional effects (Kim et al., 2012), it seems necessary to supplement the understory vegetation. In addition, since it cannot be restored to a mature forest in a short period of time, it would be desirable to restore the understory vegetation by modeling a forest dominated by evergreen broad-leaved trees (Oh and Park, 2003; Park et al., 2018), which is the middle stage of the succession series.
Conclusion
The windbreak forest created on the southwest coast is a cultural landscape that represents the identity of coastal villages, and has high importance in both tangible and intangible assets, including for village rituals held there to pray for the well-being of the village and a good catch. In addition to historical and cultural aspects, the forest has been suggested as having high value in terms of ecology and disaster prevention, but in reality it is recognized only as a tourism resource for generating profits. As this has led to the development of surrounding areas, including the installation of amenities, and the creation of accommodations and commercial facilities, the damage is being aggravated, but there are no protection and management measures. For the Yesong-ri Evergreen Forest, which was designated as Natural Monument No. 40, an attempt was made to restore the evergreen broad-leaved forest after demolishing the buildings, but this was just a one-off measure without any comprehensive plan related to the establishment of protected areas, demolition of buildings, and restoration of vegetation. To protect the windbreak forests on the west coast, it seems that based on a comprehensive plan, it is not only necessary to determine the precise state of the forest and attempt its physical restoration, but also to raise residents’ awareness of the forest by publicizing its value and the necessity of its protection. Regarding the use, it is required to propose a specific management manual that prohibits the use of trails other than walking decks after installing fences, or alternately applies a rest-year system for restoration by dividing the zones. In addition, while studies related to the value and utility of windbreaks have continuously been conducted, long-term monitoring of vegetation restoration and management effects has not been attempted. In the future, along with surveys of the status or utility of windbreak forests, it is considered that the effect of restoration should be verified.