Introduction
Plant growth is affected by environmental factors such as light, temperature, moisture, and salts. High salt concentrations in soil cause damages due to nutrient deficiency and metabolic disorder of the plant resulted from imbalance in ion concentrations that follow the increase of Na+ and ClŌłÆ ion concentrations as well as water deficit within the plant due to the difference in water potential of root zones and soils. In addition, plants show lower growth caused by the decrease in photosynthetic capacity due to damaged chloroplasts and the increase in reactive oxygen species (ROS) (Allakhverdiev et al., 2000; Singh and Prasad, 2009). The difference in salt stress tolerance varies depending on the species, and the plant generates antioxidants such as proline and alleviates stress, thereby preventing physiological damages.
The plant shows an increase in reactive oxygen species (ROS) due to all kinds of environmental stress. The constant increase of ROS becomes a major cause that inhibits overall plant production, and this ROS is eliminated by antioxidant enzymes such as superoxide dismutase (SOD), ascorbate peroxidase (APX), peroxidase (POD), and catalase (CAT) (Noctor and Foyer, 1998). SOD in particular promotes breakdown of superoxide anion radicals generated when the plant is stressed, transforming them into hydrogen peroxide and oxygen molecules, and it is an important enzyme for the early stress process (Fridovich, 1978). Moreover, proline is a type of amino acid that is known to be a typical osmoticum that controls water potential by adjusting the osmotic pressure of cells as a defense mechanism for salt stress and water of the plant, and it also turns out to affect cell wall proteins of certain plants (Nanjo et al., 1999). Proline is accumulated when plants receive salt or water stress, which increases tolerance (Delauney and Verma, 1993; Szabados and Savour├®, 2010).
Veronica L. belongs to Scrophulariaceae, and approximately 500 species are distributed in most parts of the Northern Hemisphere and some parts of the Southern Hemisphere (Albach et al., 2005). And the Veronica genus include subgenus Pseudolysimachion, there are total 15 species of plants in this subgenus of Veronica growing in Korea (KPNI, 2022). Veronica pusanensis is an endemic species growing only along the coasts of Busan (Chung et a l., 2017) that is also designated by Korea Forest Service as a critically endangered species (CR) (Korea National Arboretum, 2021). But, Y. N.Lee changed the name of V. pusanensis as Pseudolysimachion pusanensis (Y. N. Lee). Y. N. Lee (Lee, 2005). Its plant length is approximately 20 cm, and it has thick leaves and white hair. The flowers bloom in blue racemes in July ŌĆōAugust that lie low on the ground, which is why the plant is used as a ground cover plant. Studies on P. pusanensis include studies on the germination physiology of seeds (Song et al., 2019) and effects on growth and blooming depending on shading (Song et al., 2020), but not enough are on environmental resistance identifying physiological responses in various environmental conditions such as moisture, dryness, and temperature. In particular, research on propagation of P. pusanensisis important as an endemic and rare species, but since it is a plant that has sufficient ornamental value as well as utility value as a gardening material, it is necessary to create an environment for its plantation and cultivation. It is also important to study environmental resistance to verify the growth of plants in various environmental conditions. Since P. pusanensis grows along certain coasts of Busan, it is necessary to identify the change in growth and physiological mechanism of the plant depending on the salts in soil while studying environmental resistance. Therefore, this study investigates the change in growth and physiological functions of P. pusanensis that has high utility value as a gardening material and identifies the tolerance to salt stress.
Research Methods
Materials for the experiment
Pseudolysimachion pusanensis seeds used in the experiment were those gathered and carefully selected in October 2020 from the greenhouse (37┬░28ŌĆ▓45.2ŌĆ│N, 127┬░35ŌĆ▓51.4ŌĆ│E) at the Useful Plant Resources Center of Korea National Arboretum located in Yongmun-myeon, Yangpyeong-gun, Gyeonggi-do and stored in a dry place for approximately 10 months at 4┬░C. They were sowed on March 12, 2021 and transplanted in rectangular pots (11.5 ├Ś 11.5 ├Ś 10.5 cm) filled with horticultural substrate on June 2, 2021. The experiment was conducted for 4 weeks from August 1 to 30, 2021 after rooting for 2 months. The early growth of P. pusanensis showed the average plant length, plant width, leaf length, and leaf width of 7.0 ┬▒ 0.52, 19.1 ┬▒ 1.42, 4.2 ┬▒ 0.24, and 3.5 ┬▒ 0.27 cm, respectively.
Growing conditions and NaCl treatment
This study was conducted in the greenhouse (37┬░28ŌĆ▓N, 127┬░35ŌĆ▓ E) at the Useful Plant Resources Center of Korea National Arboretum, and overhead irrigation was applied collectively before starting the experiment. To induce salt stress, 400 mL/pot of NaCl was irrigated once a week in each of the 5 levels of concentration per treatment, such as 0, 50, 100, 200, and 400 mM. In addition, 400 mL/pot was irrigated through overhead irrigation once a week to keep the soil from drying. For the experimental plot, 9 individuals were completely randomized in each treatment repeatedly. Temperature and humidity in the greenhouse were measured every day using the environmental data logger (WatchDog 1000 Series Micro Station, Spectrum Tech. Inc., Plainfield, IL, USA). The mean temperature of the green-house during the experiment was 26 ┬▒ 2 ┬░C, and relative humidity was 85 ┬▒ 5%.
SOD and proline content analysis
We measured superoxide dismutase (SOD) and proline content after 3 weeks to identify the physiological responses due to salt stress of P. pusanensis. SOD activity is measured using the photochemical NBT method that can measure the ability to inhibit reduction of nitroblue tetrazolium (NBT) (Beyer and Fridovich, 1987). For the sample, 0.2 g of leaves were ground finely with a mortar inside liquid nitrogen, homogenized with 2 mL of extraction buffer (pH 7.8, 50 mM phosphate, 0.1% BSA, 0.1% ascorbate, 0.05% mercaptoethanol (W/V)), and centrifugated for 10 minutes using a centrifuge (VS-15CF, VISION Scientific Co., Ltd., Korea) at 4┬░C, 15,000 ├Ś g. The separated supernatant was mixed with 1 mL of buffer solution (50 mM phosphate, pH 7.0, 0.2 mMEDTA) and used as an enzyme activity measurement solution. The 60 ╬╝L solution extracted and 3.0 mL of assay-buffer (pH 7.8, 50 mM phosphate, 9.9 mM L-methionine, 57 ╬╝M NBT, 0.0044% Riboflavin) were mixed, and 2000 lux light source was irradiated for 10 minutes with fluorescent lamp LED, which was then measured at absorbance 560 nm using a spectrophotometer (X-ma 1200, Human Co., China).
Proline content was measured using the method by Bates et al. (1973), and leaves with the fresh weight of 0.5g were ground after adding liquid nitrogen and homogenized by adding 10 mL of 3% sulfosalicylic acid, after which they were centrifugated at 4┬░C, 4,000 ├Ś g for 10 minutes. We mixed 1 mL of acid-ninhydrin (acetic acid 60 mL + 6 M phosphoric acid 40 mL + ninhydrin 2.5 g) and 1 mL of acetic acid into 1 mL of supernatant and treated the samples for 60 minutes at 95┬░C using a constant temperature water tank (WB-11, DAIHAN Scientific Co., Ltd., Korea), after which they were cooled. We then added 3mL of toluene into the solution where the responses ended, stirred for 30 seconds, kept for 10 minutes at room temperature, and separated the color formation of the reacting solution. The absorbance was measured at 520 nm of proline using toluene as the blank.
Chlorophyll content and chlorophyll fluorescence measurement
For chlorophyll fluorescence analysis, we selected 6 leaves per treatment 4 weeks after the experiment and measured maximum quantum yield (Fv/Fm) using a chlorophyll fluorescence analyzer (Maxi Imaging-PAM-series, Heinz Walz GmbH, Effeltrich, Germany) after 30 minutes of dark adaptation.
For chlorophyll content, we used the same leaves as chlorophyll fluorescence measurement and used the method by Yoshida et al. (1971). We ground 0.5 g of leaves for each concentration treatment plot, put them in 50 mL plastic tubes with 80% acetone, extracted for 48 hours in a 4 ┬░C refrigerator, and measured the absorbance at 663 nm and 645 nm using a spectrophotometer.
Growth survey and statistical analysis
To verify the growth of P. pusanensis due to salt stress, we measured the plant length, plant width, leaf length, leaf width, number of leaves, root length, and top and root fresh weight and dry weight 4 weeks after salt treatment. We measured the length from the soil surface up to the highest part of the plant for plant length, the greatest width of the plant for plant width, and the biggest leaves for leaf length and leaf width. We also measured only the leaves that are at least 1 cm to calculate the number of leaves. Top and root fresh weight was measured using an electronic scale (Shimadzu Analytical Balance AUW2200, Shimadzu, Japan) immediately after the experiment, and dry weight was measured after drying for 72 hours in a 70 ┬░C conditioning oven.
For statistical analysis, we conducted an analysis of variance using SPSS (SPSS 23.0, IBM Corporaton, USA), and conducted a posttest at a 5% significance level using DuncanŌĆÖs Multiple Range Test when there was a significant statistical difference. The graph was drawn using Sigma Plot 10.0 (SPSS Inc., IL, USA).
Results and Discussion
Growth characteristics
As a result of verifying growth of Pseudolysimachion pusanensis depending on NaCl concentrations, there was no statistical significance in other indicators of plant growth aside from plant length, plant width, number of leaves, fresh weight, and dry weight, but the overall growth tended to be superior at 50 and 100 mM on average, and top and root fresh weight and dry weight were also high. The means of plant length, plant width, and number of leaves were highest with 8.6, 25.1 cm, and 137.1 leaves at 50 mM. Leaf length and leaf width were 4.6 and 3.7 cm, and root length was 27.6 cm (Table 1, Figs. 1A, 1B). The means of leaf length and leaf width were highest with 5.0 and 3.9 cm at 100 mM, and there was no statistical difference despite the mean difference between treatment concentrations. The means of plant length, plant width, root length, and number of leaves were lower than 50 mM with 6.9, 24.2, 26.2 cm, and 116.3 leaves. At 200 mM, top plant length, plant width, leaf length, and leaf width were 6.4, 18.2, 4.2, and 3.6 cm, and the number of leaves was 93.9. Compared to the control plot, there was no significant difference in leaf length and leaf width aside from plant length, plant width, and number of leaves. There was also no statistical significance in root zone, but compared to the control plot where the average root length was 26.6 cm, the root length at 200 mM was 23.6 cm, which is 3cm shorter. There was also a difference in root fresh weight and dry weight. This may be due to the fact that at least 200 mM of salt accumulation in the soil caused a direct damage to the roots, inhibited nutrient uptake, and slowed down the growth of the root zone. Continuous salt treatment of at least 200 mM will not only deteriorate the roots but also overall top growth, ultimately showing statistical significance. Moreover, the overall growth was lowest at 400 mM, with plant length 5.2 cm, plant width 14.9 cm, leaf length 3.8 cm, leaf width 3.3 cm, number of leaves 28.1, and root length 19.2 cm. In general, salt stress inhibits plant growth (Chartzoulakis and Klapaki, 2000), but low concentrations of salt treatment in various plants increase overall growth, whereas high concentrations of salt treatment inhibit growth. Different plant species are known to have different tolerance to salt stress (Aziz and Khan, 2001; Abdul Qados, 2011; Shim et al., 2012). P. pusanensis also showed better top and root growth in low concentrations of salt treatment such as 50 and 100 mM compared to the control plot, while also showing high fresh weight and dry weight. However, root development was inhibited from 200 mM, and top and root growth declined remarkably at 400 mM, which indicates that this plant species must be grown at salt concentrations lower than 200 mM in order to show constant growth.
SOD, proline, and MDA contents
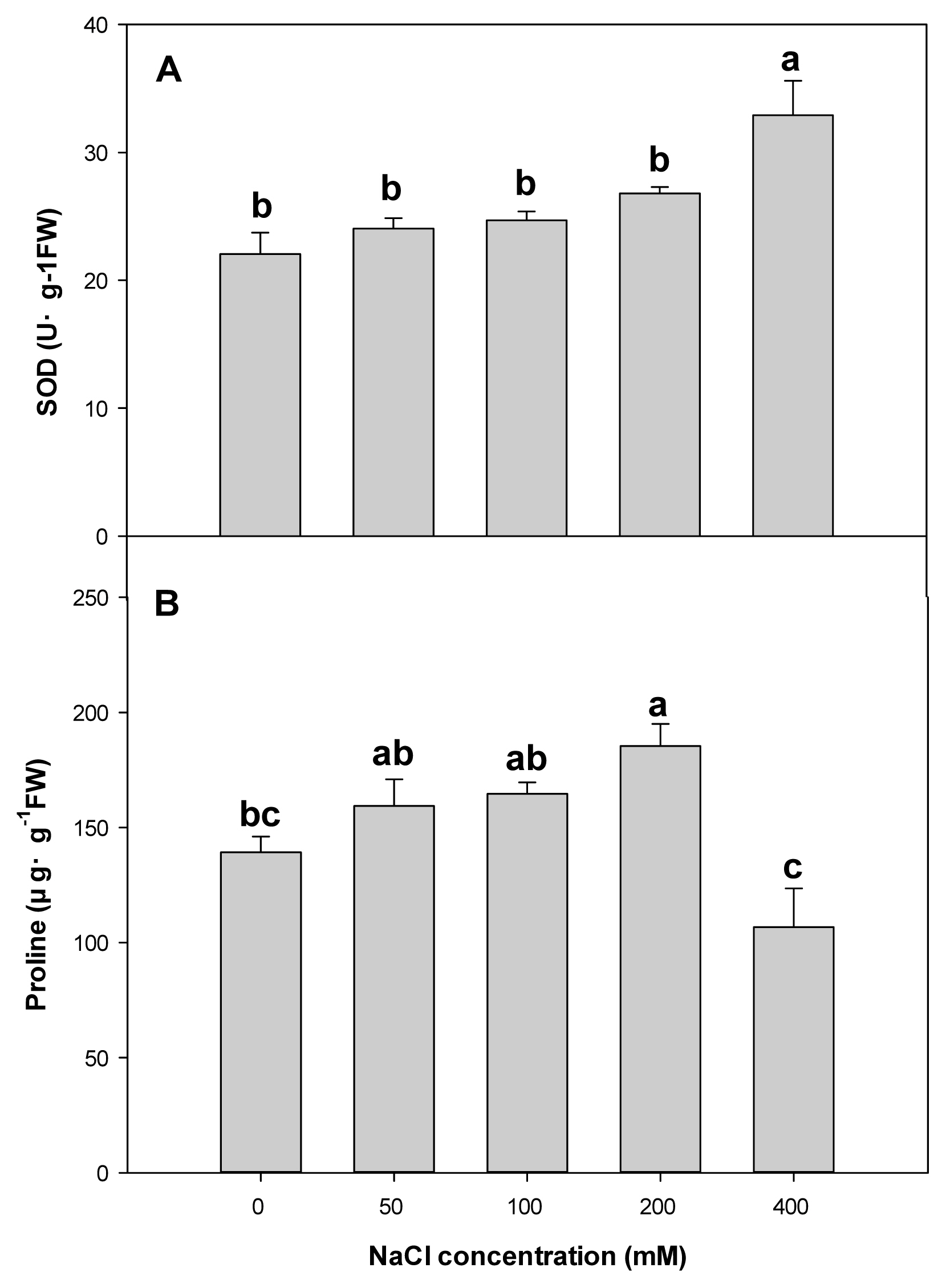
As a result of analyzing the change in SOD activity due to salt stress of P. pusanensis, the control plot showed 22.1 U┬ĘgŌłÆ1, and 50, 100, 200, and 400 mM showed 24.0, 24.7, 26.8, and 32.9 U┬ĘgŌłÆ1 respectively, proving that the average SOD activity increased compared to the control plot depending on the NaCl treatment levels. There was no statistical significance up to 200 mM, but statistical significance was found at 400 mM (Figure 2A). SOD increases to eliminate ROS formed when the plant receives environmental stress such as water and salt (Noctor and Foyer, 1998). Yang et al. (2021) discovered that SOD activity of Exochorda serratifolia increased along with CaCl2 concentrations, proving that this species receives salt stress. Aalipour et al. (2020) reported that there was an increase in SOD activity after irrigating 50% less water than the control plot of Cupressus arizonica for 5 months. P. pusanensis showed a difference in the average SOD activity among NaCl concentration levels, and considering that there was significance at 400 mM, which is a high concentration, SOD content has changed to eliminate ROS generated due to salt stress.
The proline content of P. pusanensis due to NaCl concentrations was 139.1 ╬╝g┬ĘgŌłÆ1FW in the control plot and 159.3, 164.6, and 185.4 ╬╝g┬ĘgŌłÆ1FW at 50, 100, and 200 mM, which is higher than the control plot (Fig. 2B). Proline is a type of amino acid that helps absorb water by adjusting water potential with a defense mechanism against water and salt stress of the plant (Nanjo et al., 1999), and an increase in salt concentration leads to an increase in proline content of halophytes and non-halophytes (Lee et al., 1999; Nam et al., 2007). P. pusanensis also showed an increase in proline content along with the increasing NaCl concentrations so that the proline inside the plant can adjust water potential due to the change in osmotic pressure caused by excessive NaCl in the soil. However, proline was lower (106.7 ╬╝g┬ĘgŌłÆ1FW) than the control plot at 400 mM, which is because the cells received strong salt damage.
Change in chlorophyll content and maximum quantum yield
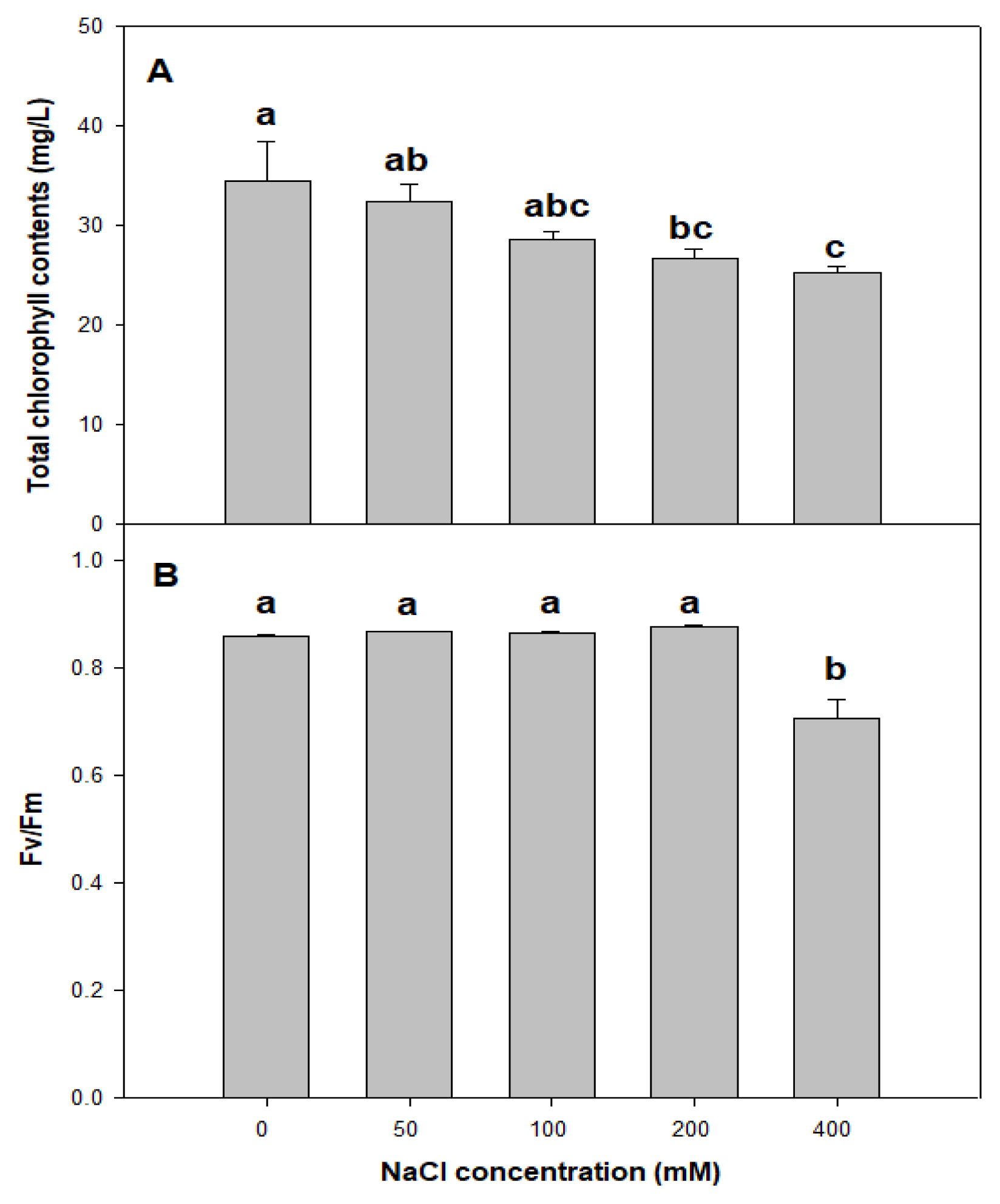
In general, increasing NaCl concentrations is known to reduce chlorophyll content by generating chlorophyllase, which is a chlorophyll breakdown enzyme (Jamil et al., 2007). Cornus kousa showed a decrease in chlorophyll content of fully mature leaves as the concentrations of CaCl2 increased (Sung et al., 2009). Lee et al. (2020) argued that an increase in NaCl concentrations leads to a decrease in chlorophyll content of Crepidiastrum sonchifolium leaves. P. pusanensis also showed the chlorophyll content of 34.6, 32.4, 28.6, 26.6, and 25.4 mg┬ĘgŌłÆ1 in the control plot and at 50, 100, 200, and 400 mM, and chlorophyll content decreased significantly as NaCl concentrations increased (Fig. 3A), which is because the increase in salt concentration promoted chlorophyll breakdown as did for Cornus kousa and Crepidiastrum sonchifolium.
Stress on plants damages the mechanism of photosynthesis, slows down photosynthesis, and reduces photosynthesis efficiency, ultimately deteriorating plant growth (Chaves and Oliveira, 2004), Chlorophyll fluorescence is a non-destructive method to study the damages from stress on the plant. Fv/Fm is used as an indicator that evaluates the maximum photosynthesis level for plant leaves through fluorescence emissions shown depending on photochemical reaction activity of the dark-adapted plant. Fv/Fm of plants that are not stressed is known to be within the range of 0.78 ŌĆō 0.84 (Yoo et al., 2012; Hazrati et al., 2016). P. pusanensis showed 0.860, 0.867, 0.861, and 0.875 in the control plot and at 50, 100, and 200 mM, which is higher than the generally known 0.84, and the maximum quantum yield decreased to 0.705 at 400 mM (Fig. 3B). According to Kim et al. (2019), Suaeda glauca, which is a halophyte, is tolerant to salts and showed an insignificant change in Fv/Fm even at high salt concentrations of 200 ŌĆō 400 mM, but Fv/Fm is known to decrease when plants sensitive to salts are stressed (Lu et al., 2002; Maricle et al., 2007). P. pusanensis did not show a significant difference in Fv/Fm from nontreatment and low concentrations (50, 100 mM) to high concentration (200 mM), but Fv/Fm was low at 400 mM because, while P. pusanensis is tolerant to salt stress up to 200 mM in terms of the mechanism of photosynthesis, the chlorophyll function is deteriorated at the high concentration of 400 mM.
In sum, as NaCl concentrations increased, SOD activity did not show a significant difference up to 200 mM but showed a significant increase at 400 mM, and proline content increased up to 200 mM and became low at 400 mM. Considering that proline content was low at 400 mM due to salt damage on cells, this result indicates that P. pusanensis receives salt stress. Moreover, considering that growth was highest at low concentrations such as 50 and 100 mM, P. pusanensis is tolerant to salts, indicating that low salt stress rather helps growth. It seems that low concentrations of salt treatment would help in plantation and cultivation of this species. Moreover, the average length of the root zone began to decrease at 200 mM, and top growth did not show a significant difference from the control plot. Considering that SOD activity and Fv/Fm did not show statistical significance in the control plot and at 200 mM, P. pusanensis seems to have strong tolerance up to 200 mM, which indicates that low concentrations of salt treatment would be suitable for creating not only growing conditions but also planting conditions.
Conclusion
This study was conducted to verify the effects of NaCl concentrations on the growth, antioxidants (superoxide dismutase, proline), maximum quantum yield, and chlorophyll content of P. pusanensis. Five levels of NaCl concentrations (0, 50, 100, 200, 400 mM) were used, and the growth was highest at 50 and 100 mM, but root growth declined before the top from 200 mM and higher. The increase in NaCl concentrations also led to the increase in SOD and proline content compared to the control plot, whereas it decreased chlorophyll content. There was no difference in the maximum quantum yield at 0, 50, 100, and 200 mM, but it decreased at 400 mM. The results proved that P. pusanensis showed an increase in SOD and proline content at 50 and 100 mM of NaCl treatment compared to the control plot, indicating that it receives salt stress, but its growth was superior at 50 and 100 mM, and there was no difference in top growth up to 200 mM compared to the control plot as well as no significant difference in maximum quantum yield, which proved that P. pusanensis has strong tolerance to salt stress.